
Artículo Inédito
Estrategias de caza y pastoreo de camélidos entre las sociedades agropastoriles (ca. siglos IV a.C. - XVI d.C.) del oeste tinogasteño (Catamarca, Argentina). Una aproximación desde los isótopos estables
Estrategias de caza y pastoreo de camélidos entre las sociedades agropastoriles (ca. siglos IV a.C. - XVI d.C.) del oeste tinogasteño (Catamarca, Argentina). Una aproximación desde los isótopos estables
Intersecciones en Antropología, vol. 21, núm. 1, 2020
Universidad Nacional del Centro de la Provincia de Buenos Aires
Recepción: 04 Abril 2019
Aprobación: 09 Abril 2019
Resumen: Este trabajo aborda las estrategias de caza y pastoreo de camélidos llevadas a cabo por las sociedades agropastoriles que habitaron el oeste tinogasteño (Catamarca) entre los siglos IV a.C. y XVI d.C. a partir del análisis de isótopos estables. Se midieron los valores de δ13C y δ15N de 48 muestras de colágeno óseo de camélidos (vicuña, llama y llama-guanaco) provenientes de distintos sitios arqueológicos del área de estudio. Se propone que las vicuñas fueron capturadas en dos sectores diferentes de la puna transicional de Chaschuil: 3500 a 3800 msnm y entre 3800 a 4300 msnm. Asimismo, se sostiene la existencia de dos estrategias de pastoreo: una, vinculada a una trashumancia de relativa larga distancia y al aprovechamiento de pasturas por encima de los 3000 msnm; y otra, relacionada con una mayor permanencia de los animales en cotas altitudinales menores (1400-3000 msnm) donde se alimentaron tanto de la vegetación silvestre como, posiblemente, de maíz. Estas estrategias presentan cambios y continuidades a lo largo del tiempo relacionadas con las variaciones ambientales y sociopolíticas del oeste tinogasteño.
Palabras clave: Camélidos, Isótopos estables, Sociedades agropastoriles.
Abstract: CAMELID HUNTING AND HERDING STRATEGIES AMONG AGROPASTORALIST SOCIETIES OF WESTERN TINOGASTA, CATAMARCA, ARGENTINA (C.400 B.C. – A.D. 1600): A STABLE ISOTOPE APPROACH. This paper addresses camelid hunting and herding strategies developed by agropastoralist societies of western Tinogasta, Catamarca, between 400 B.C. and A.D. 1600 through the analysis of stable isotopes. Isotopic δ13C and δ15N values of 48 bone collagen samples of camelid (vicuña, llama and llama-guanaco) from several archaeological sites in the study region were measured. It is proposed that vicuñas were captured in two different sectors of the transitional puna of Chaschuil, between 3500 and 3800 masl and between 3800 and 4300 masl. Likewise, we propose the existence of two herding strategies, one related to a relatively long-distance transhumance and the use of pastures above 3000 masl and the other related to longer stays in lower areas (1400-3000 masl) where the llamas consumed wild plants and, possibly, maize. These strategies present changes and continuities through time related to environmental and socio-political changes in western Tinogasta.
Keywords: Camelids, Stable isotopes, Agropastoralist societies.
INTRODUCCIÓN
Los camélidos tuvieron una significación crucial respecto del desarrollo de las sociedades agropastoriles del pasado prehispánico del Noroeste argentino. En los últimos años, los estudios de isótopos estables permitieron delinear las estrategias de caza de vicuñas y pastoreo de llamas llevadas a cabo por dichas sociedades. Los trabajos consideraron no solo valores de δ13C y δ15N de especímenes de camélidos arqueológicos sino también su contexto de recuperación, la estructura vegetacional de la región de estudio, las características ecológicas y etológicas de las especies de camélidos y valores de δ13C y δ15N de camélidos actuales. Esto posibilitó la discusión de las estrategias de movilidad y alimentación empleadas por los pastores del pasado respecto de sus llamas (Izeta et al., 2009; Yacobaccio et al., 2010; Srur et al., 2012; Dantas et al., 2014; Mengoni Goñalons, 2014; Grant, 2017) y la delimitación de potenciales áreas de captura de vicuñas (Mengoni Goñalons, 2014; Grant, 2017).
En relación con el manejo de las llamas, los estudios de isótopos estables realizados tanto en la región valliserrana como en la puneña indican dos tipos de estrategias empleadas por los pastores del pasado. La primera supone un control estricto sobre los rebaños, con una movilidad relativamente restringida y, posiblemente, con un complemento dietario de maíz (Izeta et al., 2009; Dantas et al., 2014; Grant, 2017). La segunda consiste en un mayor rango de movilidad de los rebaños, los cuales habrían consumido vegetales circundantes al sitio del cual fueran recuperados los especímenes analizados, así como también de aquellos emplazados en otros ambientes y/o cotas altitudinales (Izeta et al., 2009; Yacobaccio et al., 2010; Srur et al., 2012; Dantas et al., 2014; Mengoni Goñalons, 2014; Grant, 2017).
Respecto de la caza de vicuñas, los estudios de isótopos estables se focalizaron en discutir áreas de captura. Considerando que la puna presenta diferencias en cuanto a la proporción y abundancia de plantas C. y C. en función de la altitud (Samec, 2014; Grant, 2016) y que las vicuñas se mueven por espacios relativamente acotados, se delimitaron dos grandes áreas de captura: a) por debajo de los 3900 msnm (hasta el límite inferior de la puna), y b) por encima de los 3900 msnm (Mengoni Goñalons, 2014; Grant, 2017).
El presente trabajo es la primera aproximación desde los isótopos estables a los restos arqueofaunísticos de Camelidae del oeste tinogasteño. Se propone discutir qué tipos de estrategias de pastoreo y caza de camélidos fueron desarrolladas por las poblaciones que habitaron dicha región entre los siglos IV a.C. y XVI d.C.; además, si estas presentaron cambios y/o continuidades a lo largo del tiempo. Así, los antecedentes sintetizados en párrafos anteriores y la consideración de la compleja dinámica de ocupación del oeste tinogasteño detallada en la subsiguiente sección resultan fundamentales para abordar los datos generados.
LA REGIÓN DE ESTUDIO
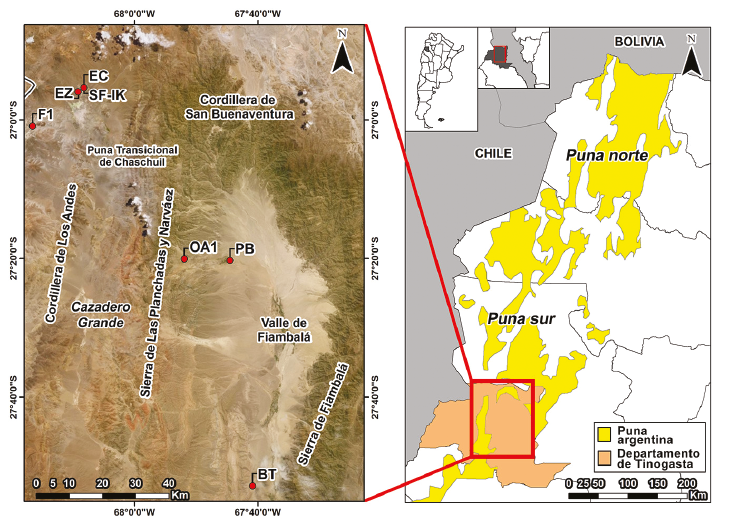
El oeste tinogasteño se localiza en el sudoeste de la provincia de Catamarca (Argentina), y abarca 13.000 km. (Figura 1). En dicha extensión se destacan el valle de Fiambalá (1400-2000 msnm); el área precordillerana, compuesta por las sierras de Las Planchadas y de Narváez y la cordillera de San Buenaventura (2200-3500 msnm); la puna transicional de Chaschuil (3500-4500 msnm) y la cordillera de los Andes (por encima de los 4500 msnm). Estos ambientes se encuentran interrelacionados mediante diversos conectores naturales que facilitan el tránsito entre ellos (Ratto, 2013).
Vervoorst (1951) definió cuatro grandes unidades vegetacionales para la región: a) estepa arbustosa tipo provincia botánica central (1400-3000 msnm, desarrollada en el valle de Fiambalá y la precordillera); b) estepa arbustosa puneña (3000-3800 msnm, asociada a la precordillera y los sectores inferiores de la puna transicional de Chaschuil); c) estepa graminosa puneña (3800-4300 msnm, en la puna transicional de Chaschuil); y d) vegas (por encima de los 3500 msnm, también en el ambiente puneño). Una primera aproximación a las vías fotosintéticas de las distintas especies sugiere un predominio de plantas C. en todas las unidades de vegetación, el cual se hace evidente a partir de los 3500-3800 msnm, tanto en la estepa graminosa puneña como en las vegas, en las que la presencia de plantas C. es sumamente escasa (Miyano, 2019).. Esta tendencia se condice con lo registrado en la puna meridional y septentrional, para donde se propuso una correlación negativa entre la altitud y la abundancia de plantas C.. Así, y al igual que en la puna transicional de Chaschuil, las especies C. se encuentran escasamente representadas por encima de los 3900 msnm (Samec, 2014; Grant, 2016). Esta información resulta relevante dado que los valores de δ13C de las plantas C. y C. presentan diferencias significativas y no se superponen; las primeras exhiben una media de -26,5‰ (con una dispersión de entre -19‰ y -35‰), y las segundas, un promedio de -12,5‰ (con una variación de entre -9‰ y -15‰) (Ambrose, 1993).
Respecto de las ocupaciones humanas, Ratto (2013) propuso el siguiente modelo de poblamiento para el oeste tinogasteño:
La ocupación del oeste tinogasteño fue un fenómeno complejo que incluyó eventos de despoblamiento y repoblamiento relacionados con la inestabilidad ambiental y la anexión al Tawantinsuyu, respectivamente. Asimismo, durante gran parte del período considerado, las poblaciones ocuparon y articularon distintos ambientes, con una interrupción entre los siglos X y XIII d.C., dada la inhabitabilidad del valle de Fiambalá. La consideración de este marco social y ambiental es imprescindible para discutir los datos generados en este trabajo.
MATERIALES Y MÉTODOS
La muestra analizada se compone de 48 especímenes óseos de Camelidae seleccionados de 12 conjuntos arqueofaunísticos los cuales provienen de siete sitios arqueológicos (Tabla 1, Figura 1).
| Sitio | Ambiente y altitud (msnm) | Conjunto | Muestras | Cronología | Referencia | |
| Años AP | Bloque temporal asignado | |||||
| Batungasta (BT) Palo Blanco (PB) | VF / 1480 | BT-E | 6 | 380±60 280±60 | XIV-XVI d.C. | Ratto (2005) |
| VF / 1900 | PB-B1-cE/F | 3 | 2210±80 | IV a.C. - X d.C. | Miyano (2018) | |
| PB-B1-cC/D | 4 | 1940±60 1790±70 | ||||
| PB-B1-cA/B | 1 | - | ||||
| PB-NH3-i/e | 2 | 1566±39 1467±39 | IV a.C. - X d.C. | Bonomo et al.(2009) | ||
| PB-NH6-i/e | 6 | 1236±37 1194±37 1190±50 | IV a.C. - X d.C. | Bonomo et al. (2010) | ||
| Ojo de Agua 1 (OA1) | PC / 2400 | OA1-R6 | 1 | 990±45 | X-XIII d.C. | Ratto (2013) |
| El Corral (EC) San Francisco Inca (SF-IK) | PT / 4000 | EC-ueII | 2 | - | X-XIII d.C. | Ratto (2013) |
| PT / 4000 | SF-IK-oPIK | 4 | 1030±60 940±80 800±70 | X-XIII d.C. | Ratto et al.(2012);Miyano et al.(2017) | |
| SF-IK-oIK | 17 | 570±60 570±50 | XIV-XVI d.C. | Miyano et al. (2017) | ||
| El Zorro (EZ) | PT / 4050 | EZ-C3-R15 | 1 | 1604±49 | IV a.C. - X d.C. | Ratto et al.(2012) |
| Fiambalá 1 (F1) | CA / 5000 | F1-R1-oF | 1 | 1499±51 1294±33 | IV a.C. - X d.C. | Ratto y Orgaz (2009) |
Batungasta (1480 msnm) y Palo Blanco (1900 msnm) se emplazan en el valle de Fiambalá. Batungasta es una instalación incaica que cuenta con diversas estructuras y dos grandes plazas (Orgaz y Ratto, 2015). Las seis muestras aquí analizadas provienen del conjunto arqueofaunístico recuperado del sector este del sitio (BT-E). Palo Blanco fue una aldea de asentamiento permanente de las primeras sociedades agropastoriles de la región. Su ocupación se extiende desde ca. 400 a.C. hasta ca. 1000 d.C., momento en que fue abandonada debido a las condiciones de inhabitabilidad del valle (Ratto et al., 2013). El sitio presenta seis núcleos habitacionales (NH) y dos basurales. Las 16 muestras aquí analizadas provienen de los conjuntos arqueofaunísticos recuperados del NH3 (PB-NH3-i/e), del NH6 (PB-NH6-i/e) y del Basural N°1 (PB-B1-cE/F, PB-B1-cC/D y PB-B1-cA/B).
Ojo de Agua 1 (2400 msnm) se emplaza en el área precordillerana y fue interpretado como un puesto de actividades pastoriles habitado de manera permanente o estacional durante la primera centuria del segundo milenio de la era (Feely y Ratto, 2009). Se seleccionó una muestra del conjunto recuperado tras la excavación del Recinto 6 (OA1-R6).
El Zorro (4050 msnm), San Francisco Inca (4000 msnm) y El Corral (4000 msnm) se localizan en la puna transicional de Chaschuil. El Zorro fue un sitio de ocupación estacional de caza y pastoreo de las sociedades agropastoriles del primer milenio de la era (Ratto et al., 2012). Del Recinto 15 del Conjunto 3 se obtuvo un conjunto arqueofaunístico (EZ-C3-R15) del cual proviene la muestra aquí analizada. El sitio San Francisco Inca presenta un patrón constructivo incaico (RPC) y cerámica imperial. Durante su ocupación se desarrollaron actividades ceremoniales patrocinadas por el Estado inca que incluyeron el consumo de alimentos y bebidas alcohólicas (Orgaz et al., 2007; Lantos et al., 2015; Miyano et al., 2017). El sitio presenta además una ocupación previa (ca. X-XIII d.C.) respecto de la cual se desconoce la disposición arquitectónica, dada su reformulación posterior siguiendo normas de construcción estatales. Se obtuvieron dos conjuntos arqueofaunísticos: uno correspondiente a la ocupación preincaica (SF-IK-oPIK) y otro a la incaica (SF-IK-oIK), de los que se seleccionaron 4 y 17 muestras, respectivamente. Finalmente, El Corral presenta dos ocupaciones: una de tiempos históricos y otra relacionada con aquella preincaica del sitio San Francisco Inca, lo que llevó a adscribirla relativamente entre los siglos X-XIII d.C. (Ratto, 2013). De esta última ocupación se recuperó un conjunto arqueofaunístico (EC-ueII), del que se seleccionaron dos muestras.
Fiambalá 1 (5000 msnm) se emplaza en la cordillera de los Andes. Fue un sitio de ocupación acotada en el tiempo donde las sociedades agropastoriles del primer milenio de la era realizaron actividades ceremoniales vinculadas a los volcanes (Ratto y Orgaz, 2009; Ratto y De Nigris, 2012). La muestra analizada proviene del conjunto arqueofaunístico recuperado del Recinto 1 (F1-R1-oF).
A partir de la consideración del modelo de poblamiento del oeste tinogasteño, cada uno de los conjuntos arqueofaunísticos fue adscripto a alguno de los tres bloques temporales propuestos por Ratto (2013): a) ca. siglos IV a.C. a X d.C.; b) ca. siglos X a XIII; y c) ca.siglos XIV a XVI d.C. (Tabla 2). Respecto del conjunto más temprano de Palo Blanco (PB-B1-cE/F), se optó por incluirlo en el primer bloque temporal, que fue extendido entonces hasta el siglo IV a.C. En relación con el conjunto PB-B1-cA/B, si bien no posee fechados radiométricos, asumimos que, por el contexto de recuperación y la posición estratigráfica, corresponde a la ocupación de la aldea durante la segunda mitad del primer milenio de la era (Miyano, 2018).
| N | Código del laboratorio | δ13C | δ15N | C:N | Tamaño | Conjunto | Bloque Temporal |
| 1 | MSR-F26 | -13,95 | 6,62 | 3,28 | LG | PB-B1-cE/F | IV a.C. a X d.C. |
| 2 | MSR-F7 | -19,37 | 4,57 | 3,32 | LG | PB-B1-cE/F | |
| 3 | MSR-F6 | -18,24 | 4,35 | 3,32 | L | PB-B1-cE/F | |
| 4 | MSR-F25 | -19,47 | 3,85 | 3,31 | LG | PB-B1-cC/D | |
| 5 | MSR-F21 | -13,76 | 2,89 | 2,85 | LG | PB-B1-cC/D | |
| 6 | MSR-F27 | -17,34 | 8,17 | 2,96 | LG | PB-B1-cC/D | |
| 7 | MSR-F24 | -14,88 | 7,44 | 2,94 | L | PB-B1-cC/D | |
| 8 | MSR-F23 | -14,92 | 4,2 | 3,35 | LG | PB-B1-cA/B | |
| 9 | MSR-606 | -17,82 | 3,8 | 3,31 | LG | PB-NH3-i/e | |
| 10 | MSR-499 | -17,9 | 3,92 | 3,32 | L | PB-NH3-i/e | |
| 11 | MSR-495 | -14,3 | 5,3 | 3,22 | LG | PB-NH6-i/e | |
| 12 | MSR-F28 | -17,69 | 6,12 | 3,28 | LG | PB-NH6-i/e | |
| 13 | MSR-F29 | -17,73 | 4,03 | 3,27 | LG | PB-NH6-i/e | |
| 14 | MSR-487 | -13,6 | 5,1 | 3,24 | L | PB-NH6-i/e | |
| 15 | MSR-497 | -17,6 | 6,3 | 3,27 | L | PB-NH6-i/e | |
| 16 | MSR-498 | -18,6 | 3,1 | 3,26 | V | PB-NH6-i/e | |
| 17 | MSR-F12 | -18,44 | 2,79 | 3,25 | V | EZ-C3-R15 | |
| 18 | MSR-604 | -18,1 | 6,8 | 3,19 | V | F1-R1-oF | |
| 19 | MSR-489 | -18,2 | 2,8 | 3,29 | LG | OA1-R6 | X a |
| 20 | MSR-F10 | -18,29 | 4,34 | 2,96 | L | SF-IK-oPIK | XIII d.C |
| 21 | MSR-F19 | -17,68 | 7,05 | 3,16 | V | SF-IK-oPIK | |
| 22 | MSR-F8 | -17,9 | 4,01 | 3,23 | V | SF-IK-oPIK | |
| 23 | MSR-F9 | -18,65 | 4,81 | 3,03 | V | SF-IK-oPIK | |
| 24 | MSR-491 | -18,7 | 3,3 | 3,21 | LG | EC-ueII | |
| 25 | MSR-605 | -17,6 | 4,3 | 3,23 | V | EC-ueII | |
| 26 | MSR-490 | -17 | 2,8 | 3,25 | L | BT-E | XIV a XVI d.C. |
| 27 | MSR-608 | -17,6 | 8 | 3,22 | L | BT-E | |
| 28 | MSR-877 | -17,55 | 7,25 | 3,59 | L | BT-E | |
| 29 | MSR-493 | -15,6 | 7,4 | 3,25 | V | BT-E | |
| 30 | MSR-600 | -16,5 | 8,1 | 3,22 | V | BT-E | |
| 31 | MSR-F20 | -17,11 | 6 | 3,17 | LG | BT-E | |
| 32 | MSR-486 | -17,3 | 3,7 | 3,19 | LG | SF-IK-oIK | |
| 33 | MSR-F5 | -17,48 | 3,58 | 3,31 | LG | SF-IK-oIK | |
| 34 | MSR-F13 | -19,02 | 4,7 | 3,24 | LG | SF-IK-oIK | |
| 35 | MSR-601 | -17,6 | 4,6 | 3,21 | L | SF-IK-oIK | |
| 36 | MSR-496 | -14 | 11,9 | 3,21 | L | SF-IK-oIK | |
| 37 | MSR-602 | -17,8 | 3,6 | 3,2 | V | SF-IK-oIK | |
| 38 | MSR-492 | -17,87 | 3,85 | 3,33 | V | SF-IK-oIK | |
| 39 | MSR-F4 | -17,35 | 4,41 | 3,37 | V | SF-IK-oIK | |
| 40 | MSR-F3 | -18,13 | 5,15 | 3,26 | V | SF-IK-oIK | |
| 41 | MSR-F15 | -18,07 | 4,36 | 3,29 | V | SF-IK-oIK | |
| 42 | MSR-F2 | -17,53 | 4,56 | 3,23 | V | SF-IK-oIK | |
| 43 | MSR-F16 | -17,88 | 5,03 | 3,3 | V | SF-IK-oIK | |
| 44 | MSR-F14 | -17,88 | 3,62 | 3,28 | V | SF-IK-oIK | |
| 45 | MSR-F17 | -18,71 | 2,67 | 3,3 | V | SF-IK-oIK | |
| 46 | MSR-F18 | -17,99 | 4,62 | 3,31 | V | SF-IK-oIK | |
| 47 | MSR-F1 | -17,73 | 3,94 | 3 | V | SF-IK-oIK | |
| 48 | MSR-F11 | -17,71 | 4,07 | 3,28 | V | SF-IK-oIK |
Los especímenes seleccionados para el análisis de isótopos estables debían cumplir con las siguientes condiciones: ser huesos con tejido denso, presentar buen estado de conservación, no encontrarse termoalterados (Yacobaccio et al., 2010; Samec et al., 2014) y estar asignados osteométricamente a alguno de los dos grupos de tamaño de camélidos: pequeño (vicuña) o grande (llama, llama-guanaco). Así, la muestra se conforma de 21 especímenes asignados a vicuña, 16 a llama-guanaco y 11 a llama2 (Tabla 2).
Para la extracción del colágeno óseo se siguió el protocolo propuesto por Coltrain y Stafford (1999). Se tomaron de 1 a 2 g de material óseo por muestra, el cual fue desmineralizado en 10 ml de 0.6M de HCl a 4°C. La solución fue cambiada cada 24 horas hasta obtener un líquido de color claro y homogéneo. El pseudomorfo de colágeno fue tratado con 0.1M de KOH por 24 horas para remover contaminantes orgánicos, ácidos húmicos y lípidos residuales, y luego se enjuagó con agua bidestilada. El pseudomorfo limpio fue congelado y liofilizado. Finalmente, unos 100 mg de colágeno liofilizado fueron gelatinizados en 5 ml de agua acificada (pH 3) por 24 horas a 120°C. El líquido resultante se filtró con una malla de 0,45 mm, se congeló y se liofilizó; de este modo se obtuvo el colágeno.
Las mediciones de las relaciones 13C/12C y 15N/14N del colágeno de las muestras se realizaron en el Laboratorio de Isótopos Estables en Ciencias Ambientales (LIECA-IANIGLA) el cual cuenta con un IRMS Thermo Delta V Advantage, interfaz universal ConFlo IV y analizador elemental Flash 2000. Las muestras estándar corresponden a Vienna Pee Dee Belemnite (VPDB) para carbono y a Ambient Inhalable Reservoir (AIR) para nitrógeno.
Los valores de δ13C de las muestras de colágeno se encuentran enriquecidos aproximadamente en un 5‰ con respecto a la dieta por el proceso de asimilación de la composición isotópica de los vegetales en dicho tejido óseo (Pate, 1994). Considerando que los camélidos son herbívoros y que los valores de δ13C de las plantas C. y C. no se superponen, las muestras de colágeno analizadas permiten inferir el aporte relativo a la dieta de dichos tipos de plantas (Ambrose, 1993), las cuales, además, presentan distribución y abundancia diferente en función de factores ambientales y altitudinales (Cabido et al., 1997, 2008; Samec, 2014; Grant, 2016).
RESULTADOS
La integridad de las señales isotópicas obtenidas fue evaluada considerando la relación atómica C:N. Todas las muestras presentan valores que se ubican dentro del rango que indica un buen estado de conservación (2,9-3,6) (De Niro, 1985). La única excepción es la muestra N°5 (2,85) (Tabla 2), por lo que se decidió descartarla.
Los camélidos de tamaño grande (llama y llama-guanaco) presentan mayor variabilidad en los valores de δ13C que los de tamaño pequeño (vicuña), puesto que cubren un rango entre -13,60 y -19,47‰ (Figura 2). Esto es esperable dado que, si se considera que la mayoría de los especímenes de tamaño grande corresponderían a Lama glama, este es un animal doméstico cuya movilidad (y alimentación) fue influenciada por los pastores. Respecto de las vicuñas, resulta esperable que exhiban valores de δ13C menos variables, debido a que su hábitat natural se encuentra restringido a ambientes puneños. Asimismo, se observa que la mayoría de los casos presentan valores de δ15N de entre 2 y 8‰, aproximadamente. Estos valores son esperables para mamíferos terrestres herbívoros en general y camélidos en particular (Grant, 2016; Samec et al., 2018).
Camélidos de tamaño pequeño (vicuña)
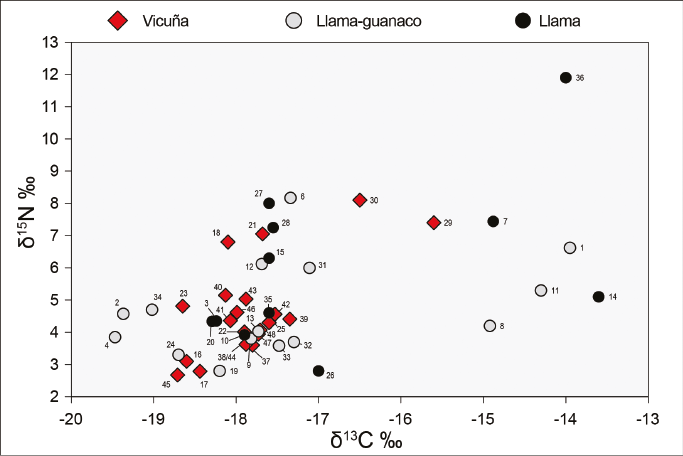
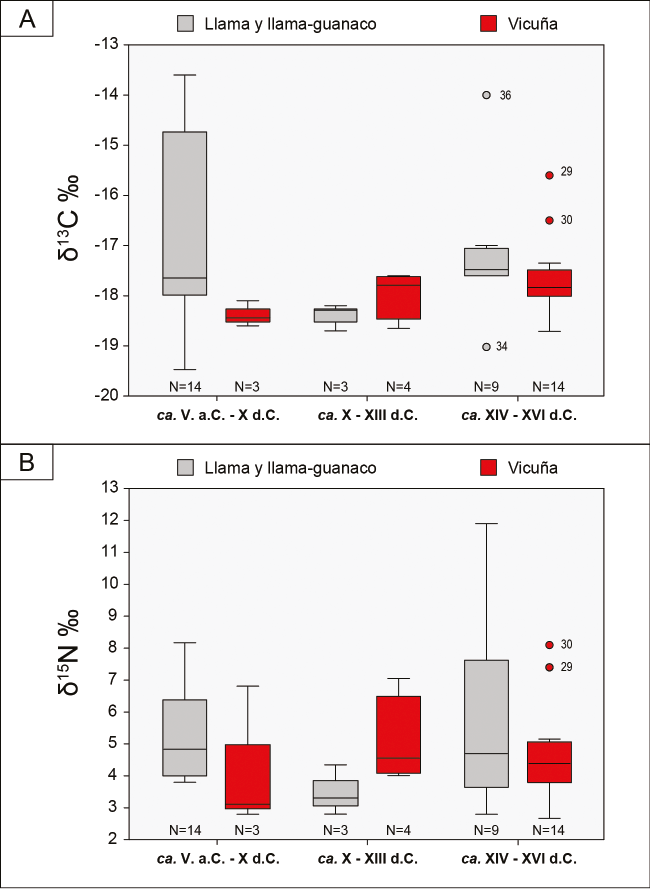
Los valores de δ13C y δ15N obtenidos de las 21 muestras se grafican en la Figura 3: los de δ13C oscilan entre -18,71 y -15,60‰, y los de δ15N, entre 2,67 y 8,10‰. Se observa que las muestras de BT-E (N°29 y 30) (Tabla 2) se distancian del resto, en tanto presentan los valores de δ13C y δ15N más enriquecidos del subconjunto (Figuras 3 y 4a y b). En relación con esto, los 21 valores de δ13C y δ15N no exhiben una distribución normal (test Shapiro-Wilk; W=0,83 .=<0,01 y W=0,89 .=<0,05, respectivamente). Sin embargo, si se excluyen las muestras de BT-E, la distribución de los 19 valores restantes es normal para δ13C y δ15N (test Shapiro-Wilk; W=0,93 .=>0,05 y W=0,91 .=>0,05, respectivamente). Así, posiblemente se trate de dos grupos de vicuñas con distintos tipos de alimentación. Aunque todos ellos se debieron alimentar principalmente de plantas C., los valores más enriquecidos de δ13C de las vicuñas de BT-E sugieren una dieta con mayor proporción de plantas C. que la de las otras o, alternativamente, una dieta con plantas C. con valores más enriquecidos.
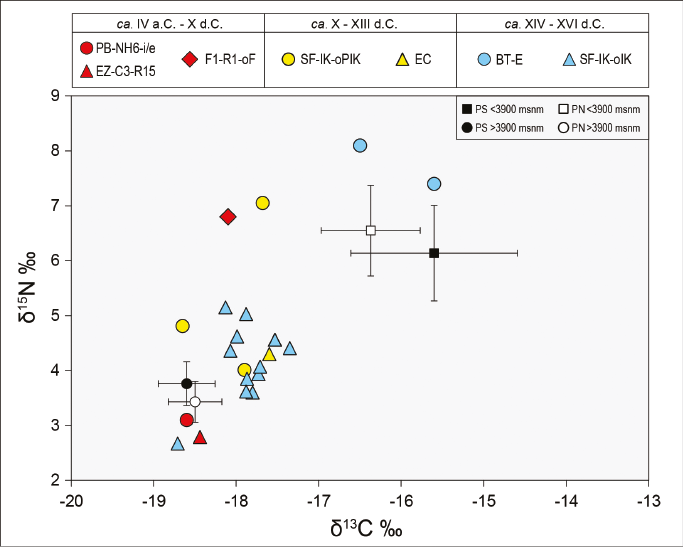
Además, en la Figura 3 se incluyen los valores de δ13C y δ15N de vicuñas actuales provenientes de la puna argentina (ver Figura 1), tanto del sector sur (PS) (Grant, 2016) como del norte (PN) (Samec et al., 2018)3. Las dos muestras de BT-E presentan valores similares a aquellos de vicuñas actuales que se alimentan en sectores inferiores a los 3900 msnm, las cuales habrían tenido un consumo mayor de plantas C.. En tanto, el resto de las muestras exhiben valores semejantes a las vicuñas actuales que se alimentan en áreas por encima de los 3900 msnm, donde la presencia de plantas C. es predominante. La diferencia señalada en la dieta de las vicuñas del pasado prehispánico del oeste tinogasteño se vincularía con su hábitat en distintos sectores de la puna transicional de Chaschuil.
Camélidos de tamaño grande (llama y llama-guanaco)
Los valores de δ13C y δ15N obtenidos de las 26 muestras se grafican en la Figura 5: los de δ13C oscilan entre -19,47 y -13,60‰, y los de δ15N, entre 2,80 y 11,90‰. Se aplicó el test Shapiro-Wilk y ni los valores de δ13C ni los de δ15N se distribuyen de manera normal (W=0,85 .=<0,01 y W=0,87 .=<0,01, respectivamente). Asimismo, se detectan dos grupos cuyos valores de δ13C se encuentran claramente separados: a) el Grupo 1 (N = 20) presenta valores entre -19,47 y -17‰; y b) el Grupo 2 (N = 6) exhibe valores más enriquecidos, entre -14,92 y -13,16‰. Esta diferencia indica una dieta disímil entre los camélidos del Grupo 1 y los del Grupo 2; estos últimos, con mayor proporción de plantas C. que los primeros. Respecto del δ15N, la mayoría de las muestras exhiben valores esperados para camélidos, que rondan entre 2,80 y 8,17‰. Sin embargo, una muestra de SF-IK-oIK (N 36) (Tabla 2) exhibe un valor de 11,90‰, y se encuentra sumamente enriquecida respecto del total de las muestras (Figuras 4b y 5).
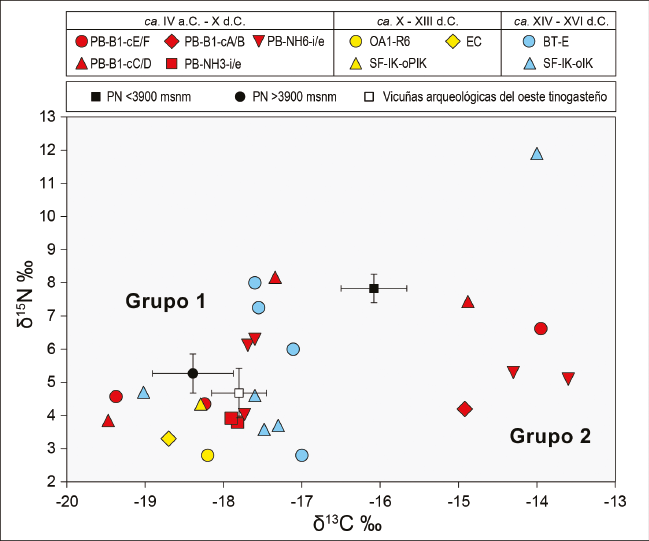
En la Figura 5 también se incluyen los valores de δ13C y δ15N de llamas actuales provenientes del sector norte de la puna argentina (PN) (Samec et al., 2018)4. y los de las 21 vicuñas arqueológicas del oeste tinogasteño presentadas anteriormente. El Grupo 1 presenta valores similares a los de las llamas actuales que se alimentan por encima de los 3900 msnm y a las vicuñas prehispánicas del oeste tinogasteño. Dicho grupo presenta muestras provenientes de distintos ambientes (valle de Fiambalá, precordillera y puna transicional de Chaschuil) y de los tres bloques temporales. El Grupo 2 –que reúne muestras provenientes de distintos momentos de la ocupación de Palo Blanco (valle de Fiambalá) y una de la ocupación incaica de San Francisco Inca (puna transicional de Chaschuil)– presenta valores de δ13C más enriquecidos que los de las llamas actuales que se alimentan entre los 3550 y 3900 msnm (Figura 5). La diferencia observada en los valores de δ13C de ambos grupos indica la existencia de dos tipos de alimentación diferentes de estos animales, por lo que, si consideramos que en su mayoría corresponden a Lama glama, estamos ante la presencia de la implementación de dos estrategias distintas en el manejo de los rebaños.
DISCUSIÓN
El análisis de los 47 valores de δ13C y δ15N permitió llevar a cabo una aproximación a la composición dietaria de los camélidos prehispánicos (IV a.C - XVI d.C.) del oeste tinogasteño, lo que propicia a su vez la discusión de las estrategias humanas vinculadas a la caza de vicuñas y al pastoreo de llamas en dicho marco espacial y temporal.
La variabilidad de los valores de δ13C y δ15N de las muestras de vicuñas y la consideración de la estructura vegetacional del oeste tinogasteño permiten aproximarnos a posibles áreas de captura. Tanto en el primer bloque temporal (IV a.C. - X d.C.) como en el segundo (X-XIII d.C.), la captura de vicuñas posiblemente se habría realizado en la estepa graminosa puneña y/o en vegas por encima de los 3800 msnm, donde predominan las plantas C.. Cabe destacar que El Zorro, El Corral y San Francisco Inca se emplazan aproximadamente a 4000 msnm, por lo que estos camélidos probablemente fueran cazados en las inmediaciones de los sitios. En tanto, las vicuñas de F1-R1-oF y de PB-NH6-i/e también deben haber sido capturadas en las cotas altitudinales más altas de la puna transicional de Chaschuil y trasladadas hacia áreas lejanas como la cordillera de los Andes (5000 msnm) y el valle de Fiambalá (1900 msnm), respectivamente. La aparición de estos especímenes en ambientes que no se corresponden con el hábitat natural de Vicugna vicugna se constituye además como evidencia del uso complementario y la articulación de los distintos ambientes por parte de las sociedades agropastoriles del primer bloque temporal. Finalmente, es posible que durante estos dos bloques temporales, la caza de vicuñas se haya dado en el marco de los movimientos de los pastores y sus llamas (Miyano, 2018).
Durante el tercer bloque temporal (XIV-XVI d.C.), las vicuñas continuaron siendo cazadas en sectores por encima de los 3800 msnm, tal como lo sugieren los valores de δ13C de las muestras de SF-IK-oIK. Recientemente se ha propuesto que, durante la ocupación incaica de San Francisco Inca, la caza de las vicuñas fue selectiva (concentrada en individuos adultos) y posiblemente se haya realizado mediante capturas colectivas en los alrededores del sitio (Miyano et al., 2017). En tanto, los valores de δ13C más enriquecidos de BT-E indican la existencia de otra área de captura, posiblemente ubicada en el límite inferior de la distribución de este animal: la estepa arbustosa puneña (3500-3800 msnm). Así, Cazadero Grande (3500 msnm) (Figura 1) se constituye como la posible área de captura de las vicuñas recuperadas en Batungasta, dado que: a) posee mayor abundancia relativa de plantas C. que los pisos altitudinales superiores (lo cual explicaría los valores de δ13C más enriquecidos de estas dos muestras); y b) presenta dos macroestructuras que fueron utilizadas durante momentos incaicos en las cacerías comunales de vicuñas mediante el chaku y/o lipi (Ratto y Orgaz, 2002-2004). De este modo, resulta posible que durante el último bloque temporal la captura de vicuñas correspondiera a eventos colectivos organizados por las autoridades estatales en distintos sectores de la puna transicional de Chaschuil (Ratto y Orgaz, 2002-2004; Miyano et al., 2017).
El análisis de los valores de δ13C y δ15N de las muestras de llama y llama-guanaco permitieron dar cuenta de la dieta de estos animales e inferir estrategias de manejo de rebaños. Se destacó que las muestras se encontraban separadas en dos grupos: uno con individuos cuyas dietas se basaron principalmente en plantas C., y otro conformado por animales con dietas de mayor aporte de plantas C4. Aquí proponemos que esta diferencia puede explicarse a partir de dos estrategias distintas de pastoreo, cuyos rangos de movilidad fueron diferentes. La primera consistió en el uso complementario de distintas áreas de pasturas (fundamentalmente, de aquellas emplazadas a 3000-4300 msnm) a partir de una trashumancia de relativa larga distancia. Así, los animales se deben haber alimentado principalmente de plantas C., en la estepa arbustosa puneña, estepa graminosa puneña y vegas. La segunda estrategia supuso una menor movilidad de los rebaños, los cuales deben haber pasado estadías más prolongadas en los sectores bajos (1400-3000 msnm) y aprovechado la vegetación de la estepa arbustosa tipo provincia botánica central (que se caracteriza por presentar la proporción más elevada de plantas C. de la región) y, posiblemente, maíz (suministrado directamente o mediante el consumo de rastrojos de la producción agrícola). Como hemos destacado, estas dos estrategias fueron propuestas en regiones ecológicamente diversas del Noroeste argentino (Izeta et al., 2009; Dantas et al., 2014; Grant, 2017).
Así, considerando los valores de δ13C y δ15N de las muestras y su cronología asociada, durante el primer bloque temporal (IV a.C.-X d.C.) y el tercero (XIV-XVI d.C.) coexistieron las dos estrategias de manejo de rebaños. En ambos momentos se produjo un movimiento de larga distancia de algunos animales mientras que otros permanecieron períodos más prolongados en pisos altitudinales menores. Así, se utilizaron y articularon los distintos ambientes del oeste tinogasteño. Respecto del posible consumo de Zea mays, cabe señalar que en toda la secuencia de ocupación de la aldea de Palo Blanco y en dos asentamientos incaicos del valle de Fiambalá (Mishma 7 y Batungasta) fueron identificados macro y microrrestos de maíz (Sempé, 1984; Ratto et al., 2010; Lantos et al., 2015).
Asimismo, durante el segundo bloque temporal (X-XIII d.C.) se desarrolló únicamente la estrategia que supone el uso complementario de las pasturas naturales ubicadas entre los 3000-4300 msnm, tanto en la zona precordillerana como en la puna transicional de Chaschuil. Cabe recordar que, durante este período, el valle de Fiambalá no presentaba condiciones ambientales que posibilitaran su habitabilidad. De este modo, las personas y sus animales se movieron exclusivamente por las tierras altas del oeste tinogasteño, donde predominan ampliamente las plantas C. por sobre las C..
CONCLUSIÓN
Los datos generados permiten sostener que las sociedades agropastoriles prehispánicas del oeste tinogasteño capturaron vicuñas en dos áreas diferentes de la puna transicional de Chaschuil y desarrollaron dos estrategias de pastoreo distintas. Por un lado, durante gran parte del período considerado (IV a.C. - XVI d.C.), las vicuñas fueron cazadas entre los 3800-4300 msnm, en la puna transicional de Chaschuil. Asimismo, durante la ocupación incaica de la región (XIV-XVI d.C.) también se utilizaron las cotas altitudinales más bajas (3500-3800 msnm) para la captura de este animal, lo cual se vincula con la organización de cacerías colectivas en distintos sectores del ambiente puneño (Ratto y Orgaz, 2002-2004). Por otro lado, en los bloques temporales IV a.C. - X d.C. y XIV-XVI d.C., los pastores utilizaron los distintos ambientes del oeste tinogasteño de manera tal que cubrieron un rango altitudinal amplio. Así, mientras algunos rebaños fueron movidos largas distancias por encima de los 3000 msnm, otros permanecieron más tiempo en los sectores bajos (1400-3000 msnm). Durante el bloque temporal X-XIII d.C., los pastores movilizaron a sus animales principalmente por encima de los 3000 msnm. Esta discontinuidad en el uso de los pisos altitudinales inferiores puede atribuirse a las condiciones ambientales desfavorables del valle de Fiambalá para su habitabilidad y tránsito durante dicho período.
El presente trabajo propició la discusión de las estrategias de caza y pastoreo llevadas a cabo por las sociedades agropastoriles que ocuparon el oeste tinogasteño entre los siglos IV a.C. y XVI d.C. Para tener un panorama más claro de las estrategias llevadas a cabo por estas sociedades resulta imprescindible complementar los análisis de isótopos estables con otras líneas de investigación, entre las que resulta fundamental la zooarqueología. Asimismo, es necesario no solo ampliar la muestra para corroborar las tendencias identificadas, sino también comenzar con la construcción de una ecología isotópica del oeste tinogasteño. Esto último ayudará a refinar las interpretaciones con una información de base que provenga exclusivamente de nuestra región de estudio.
REFERENCIAS
Ambrose, S. (1993). Isotopic analysis of paleodiets: Methodological and interpretive considerations. En M. Sandford (Ed.), Investigations of ancient human tissue. Chemical analyses in anthropology (pp. 59-130). Gordon and Breach.
Bonomo, N., Cedrina, L., Osella, A. y Ratto, N. (2009). GPR prospecting in a prehispanic village, NW Argentina. Journal Applied Geophysics, 67(1), 80-87.
Bonomo, N., Osella, A. y Ratto, N. (2010). Detecting and mapping buried buildings with GPR at an ancient village in Northwestern Argentina. Journal of Archaeological Science, 37, 3247-3255.
Cabido, M., Ateca, N., Astengiano, M. y Anton, A. (1997). Distribution of C3 and C4 grasses along an altitudinal gradient in Central Argentina. Journal of Biogeography, 24, 197-204.
Cabido, M., Pons, E., Cantero, J., Lewis, J. y Anton, A. (2008). Photosynthetic pathway variation among C. grasses along a precipitation gradient in Argentina. Journal of Biogeography, 35, 131-140.
Coltrain, J. y Stafford, T. (1999). Stable Carbon Isotopes and Salt Lake Wetlands Diet: Towards an Understanding of the Great Basin Formative. En B. Hemphill y C. Larsen (Eds.), Prehistoric Lifeways in the Great Basin Wetlands: Bioarchaeological Reconstruction and Interpretation (pp. 55-83). University of Utah Press.
Dantas, M., Figueroa, G., Laguens, A. y Izeta, A. (2014). Isótopos estables, dieta de camélidos y diferenciación social (Valle De Ambato, Catamarca, Argentina, Siglos VI-XI D.C.). Revista Chilena de Antropología, 30, 90-97.
De Niro, M. (1985). Postmortem preservation and alteration of in vivo bone collagen isotope ratios in relation to palaeodietary reconstruction. Nature, 317, 806-809.
Feely, A. y Ratto, N.(2009). Variaciones de los conjuntos cerámicos de unidades domésticas: aldeas y puestos formativos del bolsón de Fiambalá (ca. 1500-1300 AP). En N. Ratto (Comp.), Entrelazando ciencias: sociedad y ambiente antes de la conquista española (pp. 99-129). EUDEBA.
Grant, J. (2016). Isótopos estables en camélidos y vegetales modernos de Antofagasta de la Sierra: hacia una ecología isotópica de la Puna Meridional argentina. Intersecciones en Antropología, 17, 327-339.
Grant, J. (2017). Of Hunting and herding: Isotopic evidence in wild and domesticated camelids from the Southern Argentine Puna (2120-420 years BP). Journal of Archaeological Science Reports, 11, 29-37.
Izeta, A., Laguens, A., Marconetto, M. y Scattolin, M. (2009). Camelid handling in the Meridional Andes during the first millennium AD: a preliminary approach using stable isotopes. International Journal of Osteoarchaeology, 19, 204-219.
Lantos, I., Spangenberg, J., Giovannetti, M., Ratto, N. y Maier, M. (2015). Maize consumption in pre-Hispanic south-central Andes: chemical and microscopic evidence from organic residues in archaeological pottery from western Tinogasta (Catamarca, Argentina). Journal of Archaeological Science, 55, 83-99.
López, G. (2003). Pastoreo y caza de camélidos en el Temprano de la Puna de Salta: Datos osteométricos del sitio Matancillas 2. Intersecciones en Antropología, 4, 17-27.
Meadow, R. (1987). Techniques for comparing bone measurement data from small simples. Trabajo presentado en Northeastern Faunal Analysis Conference. Storrs.
Mengoni Goñalons, G. (2014). Isótopos estables en camélidos del Período Tardío del Noroeste de Argentina (NOA): Estrategias de uso y manejo de rebaños. Revista Chilena de Antropología, 30, 68-75.
Miyano, J. P. (2018). El uso de animales por las sociedades agropastoriles tempranas: análisis zooarqueológico de un basural de la aldea de Palo Blanco (valle de Fiambalá, Catamarca). Arqueología, 24(1), 77-101.
Miyano, J. P. (2019). Humanos y animales en el oeste tinogasteño (Catamarca, Argentina): su relación a lo largo del tiempo desde el siglo IV a.C. hasta la actualidad [tesis doctoral inédita, Universidad de Buenos Aires].
Miyano, J. P., Lantos, I., Ratto, N. y Orgaz, M. (2017). Animales e incas en el oeste tinogasteño (Catamarca, Argentina). Latin American Antiquity, 28(1), 28-45.
Orgaz, M., Feely, A. y Ratto, N. (2007). La cerámica como expresión de los aspectos socio-políticos, económicos y rituales de la ocupación Inka en la puna de Chaschuil y el valle de Fiambalá (Departamento Tinogasta, Catamarca, Argentina). En A. Nielsen, C. Rivolta, V. Seldes, M. Vázquez y P. Mercolli (Eds.), Procesos sociales prehispánicos en el sur andino. La vivienda, la comunidad y el territorio (pp. 237-258). Brujas.
Orgaz, M. y Ratto, N. (2015). Estrategias de ocupación incaica al sur del Tawantinsuyu (Tinogasta, Catamarca, Argentina): La apropiación de paisajes sagrados y la memoria social. Ñawpa Pacha, 35(2), 217-235.
Pate, F. (1994). Bone chemistry and paleodiet. Journal of Archaeological Method and Theory, 1, 161-209.
Ratto, N. (2005). La arqueología del Bolsón de Fiambalá a través de los Estudios de Impacto (Dpto. Tinogasta, Catamarca, Argentina). Actas das I Jornadas Internacionais Vestígios do Passado, 1-30.
Ratto, N. (2013). A modo de introducción: la articulación de estudios arqueológicos, paleoambientales e históricos en el oeste tinogasteño (Catamarca). En N. Ratto (Comp.), Delineando prácticas de la gente del pasado. Los procesos socio-históricos del oeste tinogasteño (Catamarca) (pp. 17-44). Sociedad Argentina de Antropología.
Ratto, N. y Orgaz, M. (2002-2004). La cacería en los Andes: registro material del chaku en la Puna meridional catamarqueña (Cazadero Grande, Tinogasta). Arqueología, 12, 72-102.
Ratto, N. y Orgaz, M. (2009). Poder, control y volcanes: el estado Inka en el volcán Incahuasi (Tinogasta, Catamarca). En N. Ratto (Comp.), Entrelazando ciencias: sociedad y ambiente antes de la conquista española (pp. 159-174). EUDEBA.
Ratto, N., y De Nigris, M. (2012). El consumo de camélidos (Vicugna vicugna) en un sitio ceremonial de la alta cordillera andina (Departamento Tinogasta, Catamarca, Argentina). Chungará, 44(2), 287-298.
Ratto, N., Carniglia, D. y Coll, L. (2012). Ocupación del área de “Los Seismiles” desde tiempos formativos a recientes (Departamento Tinogasta, Catamarca): nuevas evidencias. Relaciones de la Sociedad Argentina de Antropología, XXXVII(1), 207-216.
Ratto, N., Rodríguez, A. y Hershey, D. (2010). Explotación y uso de recursos vegetales en sitios arqueológicos del área cordillerana y del valle mesotérmico (Dpto. Tinogasta, Catamarca, Argentina). Actas del XVII Congreso Nacional de Arqueología Chilena, II: 1267-1278.
Ratto, N., Montero, M. C., Hongn, F. y Valero Garcés, B. (2013). La historia ambiental de las sociedades productivas del oeste tinogasteño (Catamarca), siglos I a XVI. En N. Ratto (Comp.),Delineando prácticas de la gente del pasado. Los procesos socio-históricos del oeste tinogasteño (Catamarca) (pp. 45-65). Sociedad Argentina de Antropología.
Samec, C. (2014). Ecología isotópica en la puna seca argentina: un marco de referencia para el estudio de las estrategias de pastoreo en el pasado. Cuadernos del Instituto Nacional de Antropología y Pensamiento Latinoamericano - Series Especiales, 2(1), 61-85.
Samec, C., Morales, M. y Yacobaccio, H. (2014). Exploring Human Subsistence Strategies and Environmental Change through Stable Isotopes in the Dry Puna of Argentina. International Journal of Osteoarchaeology, 24, 134-148.
Samec, C., Yacobaccio, H. y Panarello, H. (2018). Stable isotope compositions of South American camelids in the Dry Puna of Argentina: A frame of reference for the study of prehistoric herding and hunting strategies. Journal of Archaeological Science Reports, 18, 628-636.
Sempé, C. (1984). Mishma N° 7. Sitio incaico del valle de Abaucán, dto. Tinogasta-Catamarca. Revista del Museo de La Plata (NS), Antropología, 65(7), 405-438.
Srur, M., Izeta, A. y Scattolin, M. (2012). Alimentación de camélidos sudamericanos en los sitios formativos de Cardonal y Bordo Marcial (Catamarca, Argentina): Una aproximación isotópica. Archaeobios, 6(1), 5-18.
Vervoorst, F. (1951). Resultados de un viaje a la cuenca de Laguna Verde (Tinogasta-Catamarca) III. Observaciones sobre la vegetación entre Tinogasta y la cuenca. Actas de la XV Semana de Geografía, 61-67.
Yacobaccio, H., Samec, C. y Catá, M. (2010). Isótopos estables y zooarqueología de camélidos en contextos pastoriles de la puna (Jujuy, Argentina). En M. Gutiérrez, M. De Nigris, P. Fernández, M. Giardina, A. Gil, A. Izeta, G. Neme y H. Yacobaccio (Eds.), Zooarqueología a principios del siglo XX: aportes teóricos, metodológicos y casos de estudio (pp. 77-86). Ediciones del Espinillo.
Notas