
Artículo Inédito
Variaciones δ13C y δ15N en huemul (Hippocamelus bisulcus) durante el Holoceno en Cerro Casa de Piedra, Santa Cruz, Argentina. Implicancias para el estudio de su distribución pasada
Variaciones δ13C y δ15N en huemul (Hippocamelus bisulcus) durante el Holoceno en Cerro Casa de Piedra, Santa Cruz, Argentina. Implicancias para el estudio de su distribución pasada
Intersecciones en Antropología, vol. 21, núm. 1, 2020
Universidad Nacional del Centro de la Provincia de Buenos Aires
Recepción: 14 Mayo 2018
Aprobación: 18 Septiembre 2019
Resumen: El objetivo de este trabajo es estudiar la evolución de los valores δ13C y δ15N de huemul a lo largo del Holoceno en la localidad arqueológica Cerro Casa de Piedra (CCP), ubicada en la provincia de Santa Cruz. Asimismo, busca determinar la profundidad temporal de las actuales diferencias isotópicas entre huemules y guanacos de esta zona, asociadas al uso del bosque y la estepa respectivamente. De esta manera, se pone en discusión la antigua distribución del huemul y la hipótesis de una retracción de la especie al bosque. Se analizaron 20 fragmentos óseos de huemul y 18 de guanaco agrupados en Holoceno temprano (10.000-8000 años AP), medio (8000-4000 años AP) y tardío (4000-presente años AP). El huemul registró una media de δ13C -20,9‰ ± 0,7‰ y de δ15N 1,3‰ ± 1‰. El δ13C y el δ15N de huemul no exhiben variaciones significativas durante el Holoceno y no se diferencian de los especímenes modernos de la especie. Esta ausencia de variación en los valores isotópicos permite asociar al huemul con las zonas de bosque o las ecotonales en esta región de Patagonia meridional durante todo el Holoceno, lo cual resta sustento a la hipótesis del uso de ambientes esteparios por parte de este cérvido en el pasado.
Palabras clave: Zooarqueología, Holoceno, Patagonia meridional, Isótopos estables.
Abstract: δ13C AND δ15N HUEMUL (SOUTH ANDEAN DEER) VARIATIONS DURING THE HOLOCENE IN CERRO CASA DE PIEDRA, SANTA CRUZ, ARGENTINA: IMPLICATIONS FOR THE STUDY OF PAST DISTRIBUTION OF HUEMUL. The aim of this paper is to study the evolution of δ13C y δ15N huemul values throughout the Holocene at the archaeological site of Cerro Casa de Piedra (CCP), Santa Cruz province. The paper also seeks to determine the temporal depth of the current isotopic differences between huemul and guanaco in this area, associated with the use of the forest and steppe respectively. Thus, the ancient distribution of the huemul is discussed and a hypothesis presented of the retraction of the species to the forest. Twenty fragments of bone of huemul and 18 of guanaco were analyzed, grouped into the Early (10,000-8000 BP), Middle (8000-4000 BP) and Late (4000 BP to current) Holocene. The δ13C y δ15N huemul values did not record variations during the Holocene and did not differ from modern specimens of the species. Absence of variation in the isotopic values allows the huemul to be associated with either the forest or the ecotonal zones in this region of southern Patagonia during the entire Holocene, which does not support the hypothesis that steppe environments were used by this cervid in the past.
Keywords: Zooarchaeology, Holocene, Southern Patagonia, Stable isotopes.
Este trabajo está dedicado con profundo afecto a nuestra amiga, colega y mentora, Mariana De Nigris. Este artículo surgió de las charlas compartidas con ella sobre huemules e isótopos. Su curiosidad y enorme entusiasmo pronto nos contagiaron y la charla tomó forma de póster, que fue presentado en el ICAZ 2014 de San Rafael. Desde ese momento, el manuscrito fue tomando forma y lamentablemente se materializó cuando Mariana ya había partido. A pesar de la tristeza, nos alegra haber cumplido su anhelo.
INTRODUCCIÓN
En Patagonia, la zooarqueología aplicada (Lyman, 1996; Lyman y Cannon, 2004) contribuyó a comprender el impacto humano sobre poblaciones de fauna local (Scartascini y Volpedo, 2013; Zangrando et al., 2014; Nye et al., 2018); recrear la distribución de especies en el pasado (Scartascini y Volpedo, 2013; Zangrando et al., 2014; Cruz et al., 2015); inferir modificaciones en las cadenas tróficas (Vales et al., 2017) y evaluar la existencia de cambios morfológicos de los animales bajo diferentes escenarios de interacción con los seres humanos (L’Heureux, 2008). En el caso del huemul (Hippocamelus bisulcus) –ciervo endémico del bosque andino patagónico de Chile y Argentina, actualmente amenazado de extinción (Black-Decima et al., 2016)–, los aportes se enfocaron en la discusión sobre la antigua distribución de la especie (Díaz, 1990, 1993, 2000; Serret, 2001; Vila et al., 2010; Flueck y Smith-Flueck, 2012). Las hipótesis, postuladas desde la biología, están basadas en crónicas y diarios de viajeros, exploradores y antiguos pobladores desde la mitad del siglo XVI en adelante, y pueden resumirse en dos planteos. El primero argumenta que el huemul se distribuía y ocupaba en el pasado los ambientes áridos de Patagonia (Díaz, 1993, 2000; Flueck y Smith-Flueck, 2012). Esta hipótesis tiene dos variantes que difieren en el tiempo en el cual la especie se retrajo. Por un lado, la que relaciona esta retracción con presiones ejercidas durante la colonización europea y la expansión de los Estados nación de Argentina y Chile (Díaz, 1993, 2000). Por otro lado, Flueck y Smith-Flueck (2012) sostienen que dicha retracción se debió a la presión predatoria de los cazadores recolectores durante la transición Pleistoceno-Holoceno. La segunda de las hipótesis sostiene que el hábitat preferido por el huemul en el pasado era el ecotono bosque-estepa (Serret, 2001; Vila et al., 2005) y que su retracción habría sido reciente, debido al avance de las estancias y la urbanización del oeste de Patagonia continental (Serret, 2001).
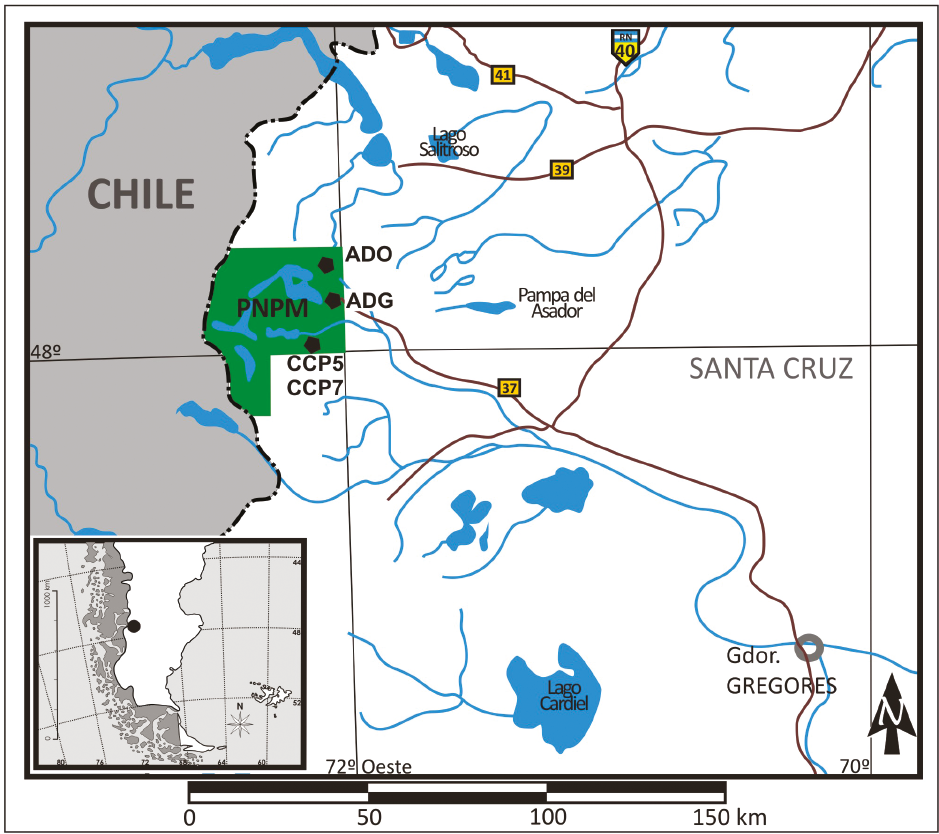
Desde la zooarqueología, a partir del estudio de los valores de los isótopos estables del carbono y del nitrógeno, se pudo establecer la relación entre el huemul y el ambiente de bosque/ecotono bosque-estepa (Barberena et al., 2011; Fernández y Tessone, 2014; Méndez et al., 2014; Tessone et al., 2014), asociación también apoyada por análisis del registro óseo de huemul recuperado en sitios arqueológicos de toda Patagonia (Barberena et al., 2011; Méndez et al., 2014; Fernández et al., 2015, 2016). Estos resultados sustentan la idea de que, en el pasado, el bosque y los espacios transicionales entre este y la estepa fueron los principales hábitats del huemul (Vila et al., 2010), en detrimento de la hipótesis que postula que la estepa fue el ambiente original de la especie (Díaz, 1990, 1993, 2000; Flueck y Smith-Flueck, 2012). No obstante, los estudios de isótopos estables también han revelado que es necesario controlar aspectos de la variación en los valores isotópicos del huemul que se expresan a escala local (Tessone et al., 2014), relacionados tanto con la heterogeneidad ambiental del área de distribución de este cérvido (Vila et al., 2010), como con la variabilidad en tiempo y espacio en su interacción con los seres humanos (Fernández et al., 2016). Así, el objetivo de este trabajo es estudiar la evolución de los valores δ13C y δ15N de huemul a lo largo del Holoceno en la localidad arqueológica Cerro Casa de Piedra (CCP), ubicada en el Parque Nacional Perito Moreno (PNPM), provincia de Santa Cruz, Argentina (Figura 1). Asimismo, se busca determinar la profundidad temporal de las actuales diferencias isotópicas entre guanacos y huemules de esta zona y, en el caso específico de esta última especie, evaluar, mediante la comparación con los registros modernos disponibles para el área (Tessone et al., 2014), cambios en los valores isotópicos del huemul producto de la colonización europea y del establecimiento del Estado nación de Argentina, uno de los principales factores de perturbación reconocido para la especie (Serret, 2001). Finalmente, con el propósito de ponderar la influencia de aspectos espaciales, se compararon los valores δ13C y δ15N de las muestras arqueológicas de ambos ungulados de CCP con los valores de guanacos esteparios del Centro Oeste de Santa Cruz (COSC) (Tessone, 2010).
ISÓTOPOS ESTABLES DEL CARBONO Y NITRÓGENO EN ARQUEOLOGÍA
Los isótopos estables del carbono y el nitrógeno medidos sobre restos óseos de origen arqueológico han sido utilizados para el estudio de dietas en el pasado. Estos han sido aplicados tanto en poblaciones humanas como en distintas especies de fauna. El fundamento de la implementación de esta metodología se basa en que los diferentes tejidos de los organismos guardan relación directa con los recursos consumidos y asimilados. En los ecosistemas, la distribución natural de los isótopos estables del carbono y nitrógeno está condicionada por diversas variables climático-ambientales, entre las que se destacan: precipitaciones, humedad relativa, temperatura, altitud, irradiación solar, el pH y los nutrientes del suelo (Tieszen, 1991).
Los isótopos estables del carbono (δ13C‰) se fijan en los ecosistemas terrestres a través de las plantas, las cuales toman el carbono atmosférico durante la fotosíntesis. Tres son las vías fotosintéticas: C., C. y CAM. Las primeras dos tienen valores δ13C contrastantes que permiten diferenciarlas, mientras que las CAM registran valores intermedios. Estas diferencias se trasladan a los siguientes niveles tróficos por medio de la discriminación isotópica. La mayoría de los árboles, arbustos y pastos pertenecen al patrón fotosintético C.. La región patagónica está dominada por plantas con vías fotosintéticas C., siendo las C. muy escasas. La distribución de valores isotópicos del carbono en plantas C. es entre -34‰ y -22‰, con un valor modal de -27‰. Este amplio rango registrado en la distribución de valores se debe a diversas variables ambientales que influyen en la síntesis del carbono por parte de la planta (Tieszen, 1991). Por ejemplo, el estrés hídrico o los ambientes salinos se manifiestan con valores δ13C elevados en las plantas. En contraposición, en los bosques lluviosos y cerrados, el decrecimiento de la irradiación solar y la poca circulación y reutilización del CO. produce valores bajos en el δ13C de las plantas. Este proceso ha sido denominado efecto del dosel vegetal (canopy effect) (Tieszen, 1991).
Por su parte, en relación con los isótopos estables del nitrógeno (δ15N‰) en el ambiente terrestre, diversos estudios evaluaron las variaciones isotópicas en especies vegetales y animales en escalas espaciales amplias, a partir de muestreos y transectas en donde existen diferencias climático-ambientales marcadas (entre otros, Murphy y Bowman, 2006; Hartman, 2011). En el ámbito de la arqueología, se destaca la discusión relacionada con el efecto de la aridez y las precipitaciones en las especies de animales y de plantas. Se han propuesto diversos modelos explicativos para este fenómeno, relacionados con el metabolismo y la fisiología de las diversas especies animales (ver síntesis en Murphy y Bowman, 2006; Hartman, 2011). No obstante, estudios posteriores concluyeron que el δ15N presente en el colágeno de las especies animales registra las variaciones de las relaciones isotópicas del nitrógeno en el suelo y las plantas, sin detectar efecto alguno relacionado con los ya citados factores metabólicos-fisiológicos (Murphy y Bowman, 2006; Hartman, 2011). Las diferencias son explicadas por una mayor apertura del ciclo del N a medida que decrecen las precipitaciones, lo cual genera mayores pérdidas en el ciclo (Hartman, 2011).
La estructuración espacial de esta variación posibilita la utilización de estos isótopos estables como marcadores geográficos en estudios ecológicos, paleoecológicos y arqueológicos. En el caso de Patagonia, las diferencias pluviométricas que se dan en sentido oeste-este repercuten en la distribución natural de los isótopos estables del carbono y del nitrógeno entre el bosque y la estepa. Estas diferencias registradas en la base de la cadena trófica se reflejan en los herbívoros (Tessone, 2010; Fernández y Tessone, 2014), de allí que huemules y guanacos modernos de Patagonia continental (Tessone et al., 2014), así como muestras arqueológicas de ambas especies (Tessone, 2010; Barberena et al., 2011; Fernández y Tessone, 2014; Méndez et al., 2014) muestren diferencias significativas en sus valores isotópicos. Por tal razón, es posible contrastar las hipótesis sobre la distribución pasada del huemul y colaborar desde la zooarqueología y los isótopos estables en la discusión sobre las políticas de conservación de la especie.
SITIOS ARQUEOLÓGICOS
El cerro Casa de Piedra es una pequeña lomada de origen volcánico emplazada en las nacientes del río Roble, cercano a la costa este del lago Burmeister, a una altura de 900 msnm, en el sector cordillerano del PNPM. En su cara norte presenta una serie de cuevas y aleros con evidencia de ocupaciones humanas entre los que se destacan los sitios Cerro Casa de Piedra Cueva 5 (CCP5) y 7 (CCP7) (Figura 1). Ambos poseen ocupaciones redundantes y continuas y manifestaciones rupestres que, en conjunto, destacan la importancia de los sitios dentro de los sistemas de movilidad de las poblaciones del área (Aschero et al., 2005). El sitio CCP5 es una cueva profunda que presenta excelentes condiciones de reparo. Es considerado un sitio de actividades múltiples ocupado entre ca. 6500 y 2500 años AP (Aschero et al., 1992). A 400 metros al oeste se ubica CCP7, también interpretado como un sitio de actividades múltiples, con una notable redundancia de ocupación y una marcada estructuración del espacio. Presenta una larga secuencia que se extiende desde ca. 10.600 años AP hasta 3400 años AP (De Nigris, 2004; Aschero et al., 2007). En ambos sitios se recuperaron abundantes restos óseos, entre los que se destacan los especímenes de guanaco (NISP 7456) y huemul (NISP 543), los cuales están presentes a lo largo de las dos secuencias de ocupación (De Nigris, 2004, 2007; De Nigris et al., 2010; Fernández, 2014).
Actualmente, el área circundante a CCP es una zona ecotonal entre el bosque de Nothofagus sp. y la estepa graminosa/arbustiva. Esta posición intermedia se habría mantenido a lo largo del Holoceno. El análisis del polen contenido en los sedimentos de los sitios arqueológicos localizados en CCP muestra que entre 10.530 ± 620 y 8920 ± 200 años AP existía en el área una estepa graminosa (Mancini, 2007). Con posterioridad a esta fecha se registra una disminución significativa en la presencia de gramíneas al mismo tiempo que aumentan Nothofagus, taxones arbustivos y plantas en cojín (Empetrum, Azorella), configuración semejante a la del actual ecotono bosque-estepa arbustiva (Mancini, 2007). El aumento de los taxones arbustivos y de Nothofagus observado ca. 6500 años AP sugiere el incremento de la temperatura estival y el aumento en la disponibilidad hídrica, señal que perdura hasta 2700 años AP (Mancini et al., 2002).
Asimismo, estudios geomorfológicos y sedimentarios dan cuenta de cambios en la configuración del paisaje circundante a CCP producto de avances y retrocesos glaciarios acontecidos en los últimos ca. 31.000 años cal. AP (Horta et al., 2017). Para el lapso de interés de este trabajo, se determinó que entre 10.169 ± 48 - 9503 ± 69 años AP (11.731-10.819 años cal. AP), los lagos Azara, Belgrano, Burmeister, Mogote, Nansen y Volcán conformaron un gran paleolago denominado Caldenius, que alcanzó una cota máxima de ca. 920 msnm, en el que CCP probablemente constituyó una península (De Nigris, 2004; Horta et al., 2017). Los registros indican que hacia 6105 ± 35 años AP (6900 años cal. AP), esa gran superficie estaba fragmentada en tres lagos, y que el cerro quedó emplazado entre el lago Burmeister y otro cuerpo de agua ubicado en el sector de las lagunas del Mie, que en ese momento tendrían una extensión mayor a la actual (Horta et al., 2017). Estos cambios en los sistemas lacustres habrían generado la disponibilidad de nuevos espacios y nichos para el establecimiento de la vegetación, lo cual impactó sobre las poblaciones animales y humanas.
MATERIALES
Las muestras fueron seleccionadas a partir de criterios cronoestratigráficos y anatómicos que aseguraron el análisis de distintos individuos. Se procesaron y analizaron 20 fragmentos óseos de huemul y 18 de guanaco. Las muestras fueron agrupadas en tres bloques temporales del Holoceno: temprano (10.000-8000 años AP), medio (8000-4000 años AP) y tardío (4000-presente años AP). Para la asignación cronológica de las muestras se utilizaron los fechados radiocarbónicos publicados (Aschero et al., 1992, 2007; De Nigris, 2004). En el caso de que una capa tuviera más de una datación, se consideró el rango entre la fecha más antigua y la más moderna, y si una de las fechas se ubicaba entre dos bloques temporales, la capa fue considerada en el bloque más representado por la dispersión de los fechados (por ejemplo, la capa 7 de CCP7 se consideró en el bloque Holoceno medio, ya que está datada entre 3920 ± 80 y 5933 ± 35 años AP). Para las comparaciones tendientes a identificar cambios a lo largo del tiempo y del espacio se utilizaron los valores publicados de muestras modernas y arqueológicas (Tessone, 2010; Tessone et al., 2014). Las primeras provienen del PNPM y las segundas conforman dos grupos de acuerdo con su procedencia: uno integrado por muestras recuperadas en sitios emplazados en la estepa del PNPM al noreste de CCP –ADO y ADG– y otro que engloba muestras recuperadas en sitios arqueológicos de la estepa del COSC –lago Cardiel, meseta de Pampa del Asador y lago Salitroso– (Figura 1). Todas las muestras son de guanaco, excepto una muestra de huemul del sitio ADG (Tessone, 2010). Las muestras de huemul y guanaco de estas diferentes escalas espaciales fueron agrupadas siguiendo los mismos criterios en los bloques temporales mencionados más arriba.
MÉTODOS
Las muestras fueron procesadas y analizadas en los laboratorios del Instituto de Geocronología y Geología Isotópica (INGEIS, CONICET-UBA). La extracción de colágeno se realizó siguiendo el protocolo utilizado en Tessone (2010). Las mediciones de las relaciones 13C/12C y 15N/14N se hicieron con un analizador elemental Carlo Erba EA1108, conectado a un espectrómetro de masas de flujo continuo Thermo Scientific Delta V Advantage a través de una interfase Thermo Scientific ConFlo IV. Los resultados son expresados como la relación del isótopo pesado sobre el liviano (13C/12C y 15N/14N) e informado como valores δ ‘‘per mil’’ (‰) en relación con estándares internacionales del carbono (VPDB) y el nitrógeno (AIR). Con el propósito de comparar muestras modernas con arqueológicas se utilizó el factor de corrección propuesto por Long et al. (2005), de acuerdo con el cual es necesario establecer el año de muerte del organismo (ver Tessone et al., 2014). Debido a los tamaños muestrales, para comparar las diferencias entre las especies y las distintas escalas espaciales se utilizó el análisis no paramétrico de Kruskal Wallis, con una significancia de p<0,05. Los resultados se expresan por medio de gráficos que promedian los valores obtenidos para cada isótopo, excepto en los conjuntos de dos o tres muestras, en que los valores se presentan de manera individual.
RESULTADOS Y DISCUSIÓN
En la Tabla 1 se registran los valores δ13C y δ15N obtenidos de las muestras de huemul y guanaco de CCP5 y CCP7, junto con los detalles osteológicos y contextuales de estas. Respecto de la preservación de los materiales analizados, los valores presentan una relación C/N comprendida en el rango normal, con una media de 3,2 ± 0,06.
| Muestra | Especie | Sitio | Cronología | Período | Hueso | Capa | δ13C‰ | δ15N‰ | C/N |
| 29584 | H. bisulcus | CCP5 | 2550 ± 90 | Holoceno tardío | Fémur | 1b | -21,3 | 0,8 | 3,2 |
| 29586 | H. bisulcus | CCP5 | 2740 ± 105 | Holoceno tardío | Falange 3 | 2 | -21,5 | 0,1 | 3,2 |
| 29589 | H. bisulcus | CCP7 | 3480 ± 70 | Holoceno tardío | Mandíbula | 1 | -21,5 | 1 | 3,3 |
| 29591 | H. bisulcus | CCP7 | 3990 ± 80 | Holoceno tardío | Maxilar | 3 | -20,3 | 3,3 | 3,4 |
| 29593 | H. bisulcus | CCP7 | 3920 ± 80 5933 ± 55 | Holoceno medio | Radio | 7 | -20,3 | 3,8 | 3,3 |
| 29585 | H. bisulcus | CCP5 | 4850 ± 110 4930 ± 160 | Holoceno medio | Metapodio | 3 | -20,3 | 1,3 | 3,2 |
| 29590 | H. bisulcus | CCP7 | 4270 ± 90 | Holoceno medio | Mandíbula | 2 | -21 | 1,5 | 3,3 |
| 29587 | H. bisulcus | CCP7 | 5120 ± 80 | Holoceno medio | Mandíbula | 4 (1) | -21,6 | 3,1 | 3,4 |
| 29600 | H. bisulcus | CCP7 | 5310 ± 110 | Holoceno medio | Mandíbula | 6 | -21,3 | 0,7 | 3,3 |
| 29588 | H. bisulcus | CCP7 | 5320 ± 90 | Holoceno medio | Mandíbula | 4 (2) | -20,6 | 1,9 | 3,2 |
| 29601 | H. bisulcus | CCP7 | 5319 ± 67 7060 ± 105 | Holoceno medio | Falange 3 | 8 | -20,8 | 1,2 | 3,3 |
| 29583 | H. bisulcus | CCP5 | 6540 ± 110 | Holoceno medio | Metapodio | 4 | -20,3 | 0,5 | 3,2 |
| 29598 | H. bisulcus | CCP7 | 7880 ± 150 | Holoceno medio | Maleolar | 11 | -22 | 1 | 3,3 |
| 29596 | H. bisulcus | CCP7 | 7905 ± 74 | Holoceno medio | Falange 1 | 13 | -21 | 0,2 | 3,2 |
| 29599 | H. bisulcus | CCP7 | 8300 ± 115 | Holoceno temprano | Metapodio | 12 | -21,7 | 0,8 | 3,3 |
| 29592 | H. bisulcus | CCP7 | 8380 ± 120 | Holoceno temprano | Falange 2 y 3 | 10 | -22 | 0,8 | 3,3 |
| 29595 | H. bisulcus | CCP7 | 8460 ± 400 | Holoceno temprano | Húmero | 14 | -19,2 | 1,4 | 3,2 |
| 29597 | H. bisulcus | CCP7 | 8920 ± 200 | Holoceno temprano | Falange 1 | 16 | -21,9 | 0,7 | 3,3 |
| 29594 | H. bisulcus | CCP7 | 9390 ± 40 | Holoceno temprano | Metapodio | 17(2) | -20,8 | 1,2 | 3,3 |
| 29602 | H. bisulcus | CCP7 | 9730 ± 100 | Holoceno temprano | Falange 3 | 15 | -20,3 | 2,2 | 3,2 |
| 33895 | L. guanicoe | CCP7 | 3480 ± 70 | Holoceno tardío | Pisciforme | 1(1) | -18,1 | 8,11 | 3,2 |
| 33428 | L. guanicoe | CCP7 | 3990 ± 80 | Holoceno tardío | Calcáneo | 3 | -19,7 | 6,2 | 3,2 |
| 33426 | L. guanicoe | CCP7 | 4270 ± 90 | Holoceno medio | Metapodio | 2 | -20 | 3,8 | 3,2 |
| 33888 | L. guanicoe | CCP7 | 3080 ± 70 6150 ± 105 | Holoceno medio | Falange 1 | 5 | -19,9 | 1,8 | 3,3 |
| 33901 | L. guanicoe | CCP7 | 5310 ± 110 | Holoceno medio | Metapodio | 6 | -19,7 | 1,53 | 3,3 |
| 33889 | L. guanicoe | CCP7 | 5320 ± 90 | Holoceno medio | Metacarpo | 4(2) | -19,9 | 2,94 | 3,2 |
| 33900 | L. guanicoe | CCP7 | 3920 ± 80 5933 ± 55 | Holoceno medio | Metapodio | 7 | -19,1 | 4,98 | 3,3 |
| 33899 | L. guanicoe | CCP7 | 5512 ± 65 | Holoceno medio | Metapodio | 9 | -21,9 | 0,01 | 3,4 |
| 33434 | L. guanicoe | CCP7 | 5319 ± 67 7060 ± 105 | Holoceno medio | Falange 2 | 8 | -22,2 | 0,4 | 3,3 |
| 33424 | L. guanicoe | CCP7 | 7880 ± 150 | Holoceno medio | Falange 1 | 11(3) | -20,1 | 1,8 | 3,2 |
| 33431 | L. guanicoe | CCP7 | 7905 ±74 | Holoceno medio | Falange 1 | 13 | -19,6 | 0,9 | 3,3 |
| 33427 | L. guanicoe | CCP7 | 8300 ± 115 | Holoceno temprano | Unciforme | 12 | -20,3 | 3,3 | 3,5 |
| 33432 | L. guanicoe | CCP7 | 8380 ± 120 | Holoceno temprano | Tibia | 10 | -20,3 | 0,5 | 3,3 |
| 33429 | L. guanicoe | CCP7 | 8460 ± 400 | Holoceno temprano | Falange 1 | 14 | -19,7 | 3,3 | 3,2 |
| 33430 | L. guanicoe | CCP7 | 8920 ± 200 | Holoceno temprano | Falange 2 | 16(2) | -20,5 | 5 | 3,3 |
| 33887 | L. guanicoe | CCP7 | 9390 ± 40 | Holoceno temprano | Metapodio | 17(4) | -19,8 | 1,57 | 3,2 |
| 33425 | L. guanicoe | CCP7 | 9530 ± 70 | Holoceno temprano | Metapodio | 18 | -20,4 | 1 | 3,2 |
| 33433 | L. guanicoe | CCP7 | 9730 ± 100 | Holoceno temprano | Metapodio | 15(2) | -19,7 | 1,9 | 3,2 |
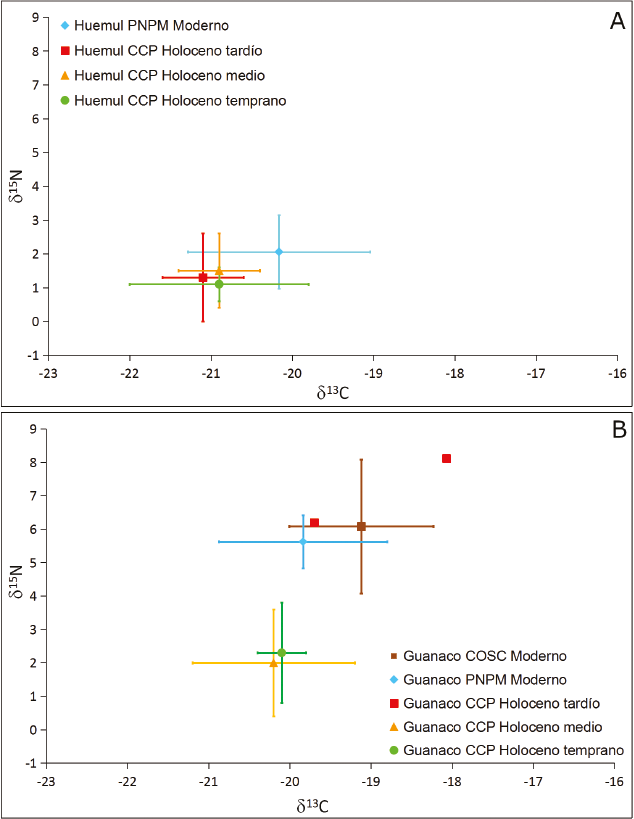
El conjunto de muestras de huemul registró una media de -20,9‰ ± 0,7‰ y de 1,3‰ ± 1‰ para el δ13C y δ15N, respectivamente. El δ13C varió entre -22‰ y -19,2‰, mientras que el δ15N, entre 0,1‰ y 3,8‰. El δ13C y el δ15N de huemul no registran variaciones durante el Holoceno, tal como se aprecia en la Figura 2A (Tabla 2) y en los resultados estadísticos (Kruskall Wallis; δ13C: p>0,05; δ15N: p>0,05). Por su parte, el guanaco presenta una media de -20‰ ± 0,9‰ y 2,7‰ ± 2,2‰ para el δ13C y δ15N. Estos valores poseen una mayor variación que los de huemul, ya que se disponen entre -22,2‰ y -18‰ para el δ13C y entre 0‰ y 8,1‰ para el δ15N (Figura 2B y Tabla 3). En términos estadísticos se registran diferencias significativas durante el Holoceno para el δ15N (Kruskall Wallis; δ15N: p<0,05), no así para el δ13C (Kruskall Wallis; δ13C: p>0,05).
| Cronología | Moderno | Holoceno tardío | Holoceno medio | Holoceno temprano | ||||
| Isótopo estable | δ13C‰ | δ 5N‰ | δ13C‰ | δ15N‰ | δ13C‰ | δ15N‰ | δ13C‰ | δ15N‰ |
| N | 7 | 7 | 4 | 4 | 10 | 10 | 6 | 6 |
| Media | -20,2 | 2,1 | -21,1 | 1,3 | -20,9 | 1,5 | -20,9 | 1,1 |
| Desvío estándar | 1,1 | 1,1 | 0,5 | 1,3 | 0,5 | 1,1 | 1,1 | 0,5 |
| Mínimo | -22,1 | 0,3 | -21,5 | 0,1 | -22 | 0,2 | -22 | 0,7 |
| Máximo | -18,6 | 3,1 | -20,3 | 3,3 | -20,3 | 3,8 | -19,2 | 2,2 |
| Cronología | Moderno | Holoceno tardío | Holoceno medio | Holoceno temprano | ||||
| Isótopo estable | δ13C‰ | δ15N‰ | δ13C‰ | δ15N‰ | δ13C‰ | δ15N‰ | δ13C‰ | δ15N‰ |
| N | 7 | 7 | 2 | 2 | 9 | 9 | 7 | 7 |
| Media | -19,8 | 5,6 | -18,8 | 7,1 | -20,2 | 2 | -20,1 | 2,3 |
| Desvío estándar | 1,0 | 0,7 | 1,1 | 1,3 | 1 | 1,6 | 0,3 | 1,5 |
| Mínimo | -20,9 | 4,2 | -19,7 | 6,2 | -22,2 | 0 | -20,5 | 0,5 |
| Máximo | -18,4 | 6,6 | -18 | 8,1 | -19,1 | 4,9 | -19,7 | 5 |
Evolución de la dieta y el uso del ambiente del huemul durante el Holoceno
Los huemules modernos poseen valores δ13C y δ15N más bajos que las muestras de guanacos modernos de las dos escalas espaciales consideradas (PNPM y COSC, Figura 3A; Tessone et al., 2014); si bien solo las diferencias en el δ15N son estadísticamente significativas (Kruskall Wallis; δ15N: p<0,05, δ13C: p >0,05). Por su parte, los guanacos de las dos regiones registran valores similares, sin diferencias significativas (δ13C y δ15N, p> 0,05; Figura 3A). Tal como se señaló, las discrepancias entre huemules y guanacos modernos reflejarían el uso de dos ambientes diferentes por parte de estos ungulados (el huemul, el bosque, y el guanaco, la estepa) y las mayores diferencias observadas en el δ15N serían producto de las variaciones existentes en la base de la cadena trófica entre estos ambientes (Tessone et al., 2014). Esto se refleja en los valores de δ13C y δ15N de vegetales modernos en distintas latitudes de Patagonia (Tessone, 2010; Fernández y Tessone, 2014).
Los resultados obtenidos en este trabajo permiten apreciar que la diferencia registrada en especímenes modernos también se observa entre los huemules de CCP y los guanacos de CCP, PNPM y COSC durante el Holoceno tardío (Figura 3B). En términos estadísticos, las diferencias entre huemules y guanacos son significativas (Kruskall Wallis; δ13C: p<0,05; δ15N: p<0,05). A su vez, los valores obtenidos en el espécimen de huemul recuperado en el sitio ADG (δ13C 22,5 δ15N 0,0; Tessone, 2010) son similares a los de los huemules de CCP, lo que refuerza las diferencias observadas entre especies durante el Holoceno tardío (Figura 3B).
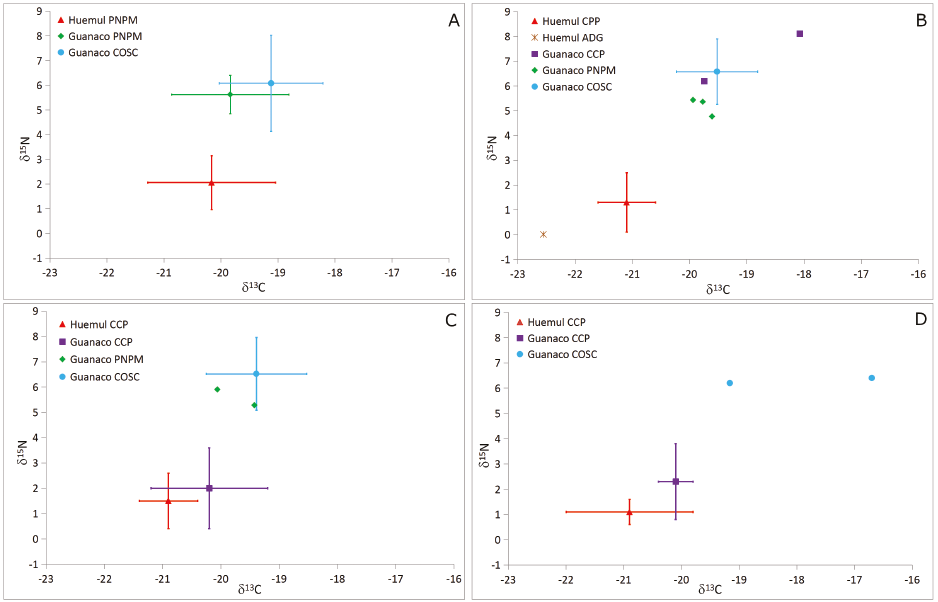
Al considerar los otros bloques temporales, el contraste entre especies presenta variaciones en relación con la escala espacial considerada. En el Holoceno medio, los valores de huemul y guanacos de CCP de ambos isótopos estables son similares y no presentan diferencias estadísticamente significativas, mientras que difieren de los valores de δ13C y δ15N de los guanacos de PNPM y COSC (Kruskall Wallis; δ13C: p<0,05; δ15N: p<0,05; Figura 3C). Cabe destacar que las muestras de guanacos del PNPM fueron recuperadas en sitios ubicados a solo 12 km al noreste de CCP, lo cual sugiere que los valores isotópicos (en especial, el δ15N) varían de manera abrupta, aun en un espacio transicional como el del PNPM. En el Holoceno temprano, las muestras de CCP exhiben el mismo patrón: valores muy similares en huemul y guanaco y diferencias marcadas con los únicos dos valores de guanaco de los que se dispone para COSC (Figura 3D).
En suma, los resultados obtenidos indican que los valores de huemul recuperados en CCP no registran diferencias en los dos isótopos a lo largo del Holoceno, y que las diferencias entre esta especie y el guanaco varían a lo largo del tiempo y en relación con la escala espacial considerada, tal como se aprecia en la Figura 4 respecto del δ15N. El aspecto más importante en relación con la evolución de la dieta y el uso del bosque es que las diferencias registradas entre los especímenes modernos de ambos ungulados se mantienen a lo largo de todo el Holoceno al comparar con las escalas PNPM y COSC, con discrepancias mayores a 1‰ y 4‰ en el δ13C y δ15N respectivamente, es decir, un nivel trófico promedio. Estas diferencias cobran mayor relevancia al considerar que las variaciones registradas en el huemul desde el Holoceno temprano hasta la actualidad son de 0,9‰ y 1,1‰ en el δ13C y δ15N respectivamente (Tabla 2). De esta manera, la ausencia de variación de los valores isotópicos del huemul a lo largo del tiempo resta sustento –al menos en lo que refiere al área del PNPM– a la hipótesis que plantea el uso de ambientes esteparios por parte de este cérvido en el pasado (Díaz, 1990, 1993, 2000; Flueck y Smith-Flueck, 2012). Como ya se mencionó, estos resultados coinciden con estudios zooarqueológicos que contemplan escalas espaciales más amplias, que sugieren que el bosque y los espacios ecotonales fueron los principales hábitats del huemul (Fernández et al., 2015, 2016).
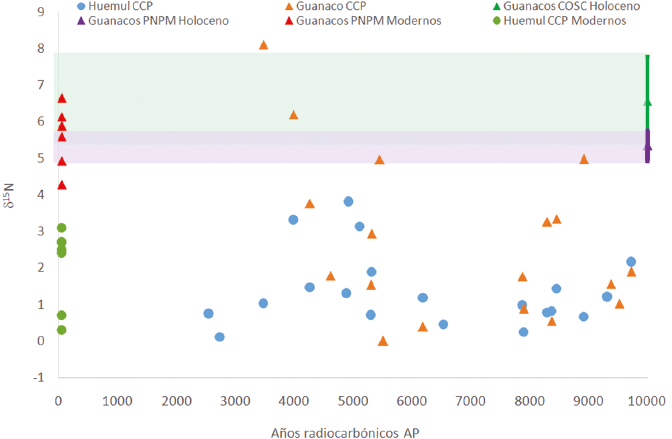
Asimismo, una lectura complementaria de estos resultados es que en el PNPM, aun en un contexto de aprovechamiento humano sostenido a lo largo de todo el Holoceno, no hubo variaciones en la señal isotópica del huemul, variaciones que podrían esperarse si la presión predatoria hubiera sido más fuerte. Destaca entonces la correspondencia entre los resultados isotópicos y los análisis zooarqueológicos efectuados en CCP (De Nigris, 2004, 2007; De Nigris et al., 2010; Fernández, 2014) y a escala regional (Fernández et al., 2015, 2016), que reafirman la idea de que el huemul siempre fue complementario o marginal a la subsistencia, salvo en contextos específicos como el interior del bosque, lo cual desestima la existencia de escenarios de sobrematanza.
Finalmente, cabe mencionar que los valores isotópicos de guanacos de CCP agrupados en los bloques temprano y medio coinciden con los patrones de alimentación del huemul (Figura 4). Los valores bajos de δ15N sugieren que estos guanacos compartían con los huemules la misma señal isotópica en la base de la cadena trófica, lo que supone que ambos ungulados se habrían alimentado en el bosque/ecotono bosque-estepa. En la costa sur de Tierra del Fuego, donde los guanacos hacen uso del bosque, Kochi et al. (2017) reportaron muestras arqueológicas con valores de nitrógeno similares a las observadas en CCP. No obstante, resulta inesperado para el COSC, ya que la gran cantidad de valores de guanaco disponibles para esta región muestra dietas más enriquecidas en 15N, asociadas a la estepa patagónica (Tessone, 2010). En efecto, las dos muestras de guanaco asignadas al bloque tardío de CCP poseen valores de d15N similares a los de COSC. De confirmarse esta tendencia con más muestras, las diferencias entre el bloque tardío y los bloques temprano y medio podrían estar relacionadas con los cambios en las cuencas lacustres del PNPM con el correr del tiempo. Estas cuencas alcanzaron su configuración moderna con posterioridad al Holoceno medio (Horta et al., 2017), lo que permite suponer variaciones en la disponibilidad de espacios y de la vegetación antes de ese momento, que afectaron la circulación y distribución de los guanacos y, a su turno, en la dieta y los valores δ13C y δ15N de este ungulado.
CONSIDERACIONES FINALES
La premisa de este trabajo es que el estudio del δ13C y δ15N en restos de huemul de sitios arqueológicos permite comprender su distribución pasada y los cambios de hábitat asociados a la interacción con las poblaciones humanas. Debido a las diferencias en las variables climáticas ambientales del bosque y la estepa y su incidencia en los ciclos del carbono y del nitrógeno en la base de las cadenas tróficas, la dieta de los herbívoros permite evaluar la distribución pasada de la especie y sus cambios a lo largo del tiempo. Las muestras de la localidad arqueológica de CCP brindan la posibilidad de controlar la existencia de variaciones en la dieta del huemul en este espacio transicional entre el bosque y la estepa en el PNPM. A esta escala espacial, la ausencia de valores isotópicos que puedan asociarse con dietas basadas en plantas que crecen en la estepa debilita la idea de que el huemul hacía uso de dicho ambiente en el pasado. Asimismo, la ausencia de variaciones en la señal isotópica de la especie, a pesar de su aprovechamiento a lo largo de todo el Holoceno, sugiere una baja presión predatoria, en consonancia con los estudios zooarqueológicos a distintas escalas espaciales (De Nigris, 2004, 2007; De Nigris et al., 2010; Fernández, 2014; Fernández et al., 2015, 2016). Por último, queda abierta la posibilidad de que las diferencias observadas en los valores isotópicos de los guanacos de CCP del Holoceno tardío y los del Holoceno temprano y medio respondan a los cambios experimentados por las cuencas lacustres del PNPM a lo largo del tiempo. Esto deberá ser evaluado por medio del análisis de un mayor número de muestras.
Agradecimientos
Nuestro agradecimiento a los profesionales y técnicos del INGEIS que colaboraron en el análisis de las muestras. Asimismo, queremos agradecer a Ana Forlano por su ayuda en la edición de las figuras. Los análisis de isótopos estables se hicieron en el marco del proyecto PICT 2013-1335, 2017-2999. Se agradece especialmente a los evaluadores anónimos, que mejoraron la versión final de este trabajo.
REFERENCIAS
Aschero, C. A., C. Bellelli, Civalero de Biset, M. T., Goñi, R., Guráieb, A. G. y Molinari, R. (1992). Cronología y tecnología en el Parque Nacional Perito Moreno (PNPM): ¿Continuidad o reemplazos? Arqueología, 2, 107-134.
Aschero, C. A., Bellelli, C., Civalero de Biset, M. T., Goñi, R., Guráieb, A. G., Molinari, R. y Espinosa, S. (2005). Holocenic Park: Arqueología del Parque Nacional Perito Moreno. Anales de la Administración de Parques Nacionales. Naturaleza y Cultura, 17, 71-119.
Aschero, C. A., Bozzuto, D., Civalero, M. T., De Nigris, M., Di Vruno, A., Dolce, V., Fernández, N., González, L. y Sacchi, M. (2007). Nuevas evidencias sobre las ocupaciones tempranas en Cerro Casa de Piedra 7. En F. Morello, M. Martinic, A. Prieto y G. Bahamonde (Eds.), Arqueología de Fuego-Patagonia. Levantando piedras, desenterrando huesos… y develando arcanos (pp. 569-576). Centro de Estudios del Cuaternario Fuego, Patagonia y Antártica (CEQUA).
Barberena, R., Méndez, C., Mena F. y Reyes O. (2011). Endangered species, archaeology, and stable isotopes: huemul (Hippocamelus bisulcus) isotopic ecology in central-western Patagonia (South America). Journal of Archaeological Science, 38, 2313-2323.
Black Decima, P. A., Corti, P., Díaz, N., Fernández, R., Geist, V., Gill, R., Gizejewski, Z., Jiménez, J., Pastore, H., Saucedo, C. y Wittmer, H. (2016). Hippocamelus bisulcus. The IUCN Red List of Threatened Species 2016. https://dx.doi.org/10.2305/IUCN.UK.2016-1.RLTS.T10054A22158895.en
Cruz, I., Muñoz, A. S., Ercolano, B., Lemaire, C. R., Pretto, A. L., Nauto, G. y Moreno, C. (2015). Apostaderos de pinnípedos en Punta Entrada (Santa Cruz, Patagonia Argentina). Explotación Humana e Historia Natural. Magallania, 43(1), 291-308.
De Nigris, M. E. (2004). El consumo en grupos cazadores recolectores. Un ejemplo zooarqueológico de Patagonia meridional. Sociedad Argentina de Antropología.
De Nigris, M. E. (2007). Nuevos datos, viejas colecciones: los conjuntos óseos de Cerro Casa de Piedra Cueva 5 (Parque Nacional Perito Moreno, Santa Cruz). Intersecciones en Antropología, 8, 253-264.
De Nigris, M. E., Lacrouts, A. y Tecce, S. (2010). Nuevos datos zooarqueológicos de las ocupaciones tempranas de Cerro Casa de Piedra Cueva 7 (Santa Cruz, Argentina). En M. A. Gutiérrez, M. De Nigris, P. M. Fernández, M. Giardina, A. F. Gil, A. Izeta, G. Neme y H. D. Yacobaccio (Eds.), Zooarqueología a principios del siglo XXI: aportes teóricos, metodológicos y casos de estudio (pp. 209-217). Ediciones del Espinillo.
Díaz, N. I. (1990). El huemul: Antecedentes históricos. Edipubli.
Díaz, N. I. (1993). Changes in the range distribution of Hippocamelus bisulcus in Patagonia. Zeitschrift für Säugetierkunde, 58, 344-351.
Díaz, N. I. (2000). El huemul (Hippocamelus bisulcus Molina, 1782): Una perspectiva histórica. En N. I. Díaz, J. A. Smith-Flueck (Eds.), El huemul patagónico. Un misterioso cérvido al borde de la extinción (pp. 1-32). L.O.L.A.
Fernández, N. L. (2014). Perfiles de mortalidad en conjuntos arqueofaunísticos holocénicos del Parque Nacional Perito Moreno (Santa Cruz, Argentina): Análisis del sitio Cerro Casa de Piedra 7 [tesis de licenciatura inédita, Universidad de Buenos Aires].
Fernández, P. M. y Tessone, A. (2014). Modos de ocupación del bosque patagónico de la vertiente oriental de los andes: aportes desde la ecología isotópica. Revista Chilena de Antropología, 30, 83-89.
Fernández, P. M., Cruz, I., Belardi, J. B., De Nigris, M. y Muñoz, S. A. (2015). Human predation and natural history of huemul (CERVIDAE; Hippocamelus bisulcus Molina) in Patagonia: A zooarchaeological Analysis. Journal of Ethnobiology, 35(3), 472-498.
Fernández, M. P., Cruz, I., Belardi, J. B., De Nigris, M. y Muñoz, S. A. (2016). La explotación de huemul (Hippocamelus bisulcus, Molina 1782) en la Patagonia a lo largo del Holoceno. Magallania, 44(1), 187-209.
Flueck, W. T. y Smith-Flueck, J. O. (2012). Huemul heresies: beliefs in search of supporting data. 1. Historical and zooarchaeological considerations. Animal Production Science, 52(8), 685-693.
Hartman, G. (2011). Are elevated δ15N values in herbivores in hot and arid environments caused by diet or animal physiology? Functional Ecology, 25, 122-131. 10.1111/ j.1365-2435.2010.01782.x
Horta, L. R., Georgieff, S. M., Aschero, C. A. y Goñi, R. A. (2017). Paleolacustrine records from Late Pleistocene e Holocene in the Perito Moreno National Park, Argentinian Patagonian Andes. Quaternary International, 436, 8-15.
Kochi, S., Pérez, S. A., Tessone, A., Ugan, A., Tafuri, M. A., Nye, J., Tivoli, A. y Zangrando, A. F. J. (2017). δ1.C and δ15N variations in terrestrial and marine foodwebs of Beagle Channel in the Holocene. Implications for human paleodietary reconstructions. Journal of Archaeological Science Reports, 18, 696-707.
L’Heureux, G. L. (2008). El estudio arqueológico del proceso coevolutivo entre las poblaciones humanas y las poblaciones de guanacos en Patagonia Meridional y Norte de Tierra del Fuego. British Archaeological Reports International Series 1751. Archaeopress.
Long, E. S., Sweitzer, R. A., Diefenbach, D. R. y Ben-David, M. (2005). Controlling for anthropogenically induced atmospheric variation in stable carbon isotope studies. Oecologia, 146, 148-156.
Lyman, R. L. (1996). Applied zooarchaeology: the relevance of faunal analysis to wildlife management. World Archaeology, 28, 110-125.
Lyman, R. L. y Cannon, K. P. (Eds.) (2004). Zooarchaeology and Conservation Biology. The University of Utah Press.
Mancini, M. V. (2007). Cambios paleoambientales en el ecotono bosque-estepa: Análisis polínico del sitio Cerro Casa de Piedra 7, Santa Cruz (Argentina). En F. Morello, M. Martinic, A. Prieto, y G. Bahamonde (Eds.), Arqueología de Fuego Patagonia. Levantando piedras, desenterrando huesos y... develando arcanos (pp. 89-94). CEQUA.
Mancini, M. V., Páez, M. y Prieto, A. (2002). Cambios paleoambientales durante los últimos 7.000 14C años en el ecotono bosque-estepa, 47-48ºS, Santa Cruz, Argentina. Ameghiniana,39(2),151-162.
Méndez, C., Barberena, R., Reyes, O. y Nuevo Delaunay, A. (2014). Isotopic Ecology and Human Diets in the Forest–Steppe Ecotone, Aisén Region, Central-Western Patagonia, Chile. International Journal of Osteoarchaeology, 24, 187-201. 10.1002/oa.2337
Murphy, B. P. y Bowman, D. M. J. S. (2006). Kangaroo metabolism does not cause the relationship between bone collagen d15N and water availability. Functional Ecology, 20, 1062-1069.
Nye, J., Zangrando, A. F., Martinoli, M. P., Vázquez, M. y Fogel, M. (2018). Cumulative Human Impacts on Pinnipeds Over the Last 7,500 Years in Southern South America. The SAA Archaeological Record, 18(4), 47-52.
Scartascini, F. y Volpedo, A. (2013). White croaker (Micropogonias furnieri) paleodistribution in the Southwestern Atlantic Ocean. An archaeological perspective. Journal of Archaeological Science, 40(2), 1059-1066.
Serret, A. (2001). El huemul. Fantasma de la Patagonia. La historia natural del ciervo andino de la Patagonia. Zaguier & Urruty Publications.
Tessone, A. (2010). Arqueología y Ecología Isotópica. Estudio de isótopos estables de restos humanos del Holoceno tardío en Patagonia meridional [tesis doctoral inédita, Universidad de Buenos Aires].
Tessone, A., Fernández, P. M., Bellelli, C. y Panarello, H. O. (2014). δ13C and δ15N characterization of modern Huemul (Hippocamelus bisulcus) from the Patagonian Andean Forest. Scope and limitations of their use as a geographical marker. International Journal of Osteoarchaeology, 24, 219-230.
Tieszen, L. L. (1991). Natural variations in the carbon isotope values of plants: implications for archaeology, ecology, and paleoecology. Journal of Archaeological Science, 18, 227-248.
Vales, D. G., Cardona, L., Zangrando, A. F., Borella, F., Saporiti, F., Goodall, R. N. P., Rosa de Oliveira, L. y Crespo, E. A. (2017). Holocene changes in the trophic ecology of an apex marine predator in the South Atlantic Ocean. Oecologia, 183(2): 555-570. https://doi.org/10.1007/s00442-016-3781-4
Vila, A. R., Díaz, N. I., Pastore, H., López R., y Faúndez, R. (2005). Distribución pasada y actual del huemul: escenarios de cambio potencial. Wildlife Conservation Society, Codeff y Publicación Técnica de la Fundación Vida Silvestre Argentina.
Vila, A., Saucedo, C. E., Aldridge, D., Ramilo, E. y Corti, P. (2010). South Andean huemul Hippocamelus bisulcus (Molina 1782). En D. J. M. Barbanti y S. González (Eds.), Neotropical Cervidology: Biology and Medicine of Latin American Deer (pp. 89-101). International Union for the Conservation of Nature; Fundação de Apoio a Pesquisa (FUNEP).
Zangrando, A. F. J., Panarello, H. O. y Piana, E. L. (2014). Zooarchaeological and Stable Isotopic Assessments on Pinniped-Human Relations in the Beagle Channel (Tierra del Fuego, Southern South America). International Journal of Osteoarchaeology, 24, 231-244.